一种提高植物抗镍性能的pgireg1s和alatp-prts双基因组及其应用
技术领域
1.本发明涉及基因工程领域,具体而言,涉及一种提高植物抗镍性能的pgireg1s和alatp-prts双基因组及其应用。
背景技术:
2.随着工业化生产和中国城市化进程不断加快,现阶段中国对采矿、制革、冶炼、电镀、烧碱制造、垃圾焚烧、污水灌溉等行业大力发展,但由于在此过程中存在的管理制度不完善,技术相对比较落后,高效循环利用效率低等状况,从而导致了大量的重金属如铅、汞、镉、钴等一大批重金属污染物进入大气、水、土壤引起了严重的环境污染问题。重金属污染不仅会导致土壤生产能力的下降,而且还可以通过根部的吸收,迁移转化到农作物根茎叶及果实中去,经过食物链最后累积到人的体内,从而危害到人的身体健康。重金属的防治和修复备受当今国际研究者所关注。
3.近年来,随着金属矿产的开采和冶炼、含镍农药和化肥的过量使用、污水农用等情况的出现,使部分地区的土壤镍含量达到原始土壤背景值得几倍甚至几十倍,远远超出土壤环境容纳能力,对我国农作物安全生产与居民健康造成严重威胁。据2014环保部与国土资源部调查显示,土壤耕地镍的点位超标率为4.8%,仅次于镉(7%)。因此,镍金属污染问题已十分严峻,引起社会各界的普遍关注,如何改善和提高我国农业生态环境质量,促进农业优质高产,保障农产品安全和人类健康,已成为当今我国农业发展面临的重大问题。
4.如何降低土壤环境中镍含量,减少对农作物产品的污染,保障生态系统尤其是人类健康已成为土壤植物营养与生态环境交叉领域的国际研究前沿热点和难点。由于土壤中重金属元素不能为土壤中微生物所降解,给污染治理增加了很大的困难,因此,如何有效的将土壤中重金属元素彻底的清除将是研究的难点。常规的土壤重金属污染治理方法,如物理方法、化学方法,由于其对技术的要求比较苛刻,另外其经济成本也比较高,而且容易造成对土壤的二次污染以及修复的过程当中可能会对土壤的结构造成破坏等原因,种种不利因素限制了其大规模的推广应用。植物修复因其修复成本低廉、易操作、不会破坏土壤结构等众多的优点,现已发展为目前土壤重金属修复最有发展前景的技术,为土壤修复开辟了新的途径。
技术实现要素:
5.本发明的目的在于提供一种提高植物抗镍性能的pgireg1s和alatp-prts双基因组及其应用,解决背景所提出的至少一个技术问题。
6.第一方面,本发明在于提供一种提高植物抗镍性能的pgireg1s和alatp-prts双基因组,所述双基因组包括:
7.膜铁转运蛋白基因pgireg1s的核苷酸序列如seq id no 1所示;
8.atp磷酸核糖转移酶基因alatp-prts的核苷酸序列如seq id no 2所示。
9.第二方面,本发明在于提供一种提高植物抗镍性能的pgireg1s和alatp-prts双基因表达载体,所述双基因表达载体为上述的双基因组与含有双35s启动子1301质粒连接构建而成。
10.第三方面,本发明还提供了上述双基因表达载体在转基因植物中的应用。
11.优先的,上述在转基因植物中的应用为将上述双基因表达载体转入转化禾本科植物,使其表达对镍的抗性和富集。
12.优先的,所述禾本科植物为水稻。
13.优先的,所述转化为农杆菌介导转化。
14.技术效果:
15.本发明采用基因合成法合成了一种来源于九节属psychotria gabriellae的pgireg1s基因和来源于镍超积累型香雪球的atp磷酸核糖转移酶alatp-prts基因,该两个基因可成功转化到水稻中,并在水稻中高效表达,得到转基因水稻;转入pgireg1s和alatp-prts基因的水稻植株和野生型水稻植株在镍抗性上有明显的差异,转基因水稻与野生型水稻相比,具有明显的抗和富集镍的能力,表明pgireg1s和alatp-prts基因的转入提高了水稻植株抗和富集镍的能力。
附图说明
16.为了更清楚地说明本发明实施例的技术方案,下面将对实施例中所需要使用的附图作简单地介绍,应当理解,以下附图仅示出了本发明的某些实施例,因此不应被看作是对范围的限定,对于本领域普通技术人员来讲,在不付出创造性劳动的前提下,还可以根据这些附图获得其他相关的附图。
17.图1为本发明含有目的基因的植物表达载体构建示意图;
18.图2为本发明实施例4中转基因水稻阳性苗的pcr扩增结果图;
19.图3为本发明实施例5中浇灌镍后转pgireg1s和alatp-prts基因水稻与野生型水稻的抗镍表型图。
具体实施方式
20.为使本发明实施例的目的、技术方案和优点更加清楚,下面将对本发明实施例中的技术方案进行清楚、完整地描述。实施例中未注明具体条件者,按照常规条件或制造商建议的条件进行。所用试剂或仪器未注明生产厂商者,均为可以通过市售购买获得的常规产品。
21.以下结合实施例对本发明的特征和性能作进一步的详细描述。
22.本发明实施例:
23.第一方面,在于提供一种提高植物抗镍性能的pgireg1s和alatp-prts双基因组,所述双基因组包括:
24.膜铁转运蛋白基因pgireg1s的核苷酸序列如seq id no 1所示;
25.atp磷酸核糖转移酶基因alatp-prts的核苷酸序列如seq id no 2所示。
26.第二方面,在于提供一种提高植物抗镍性能的pgireg1s和alatp-prts双基因表达载体,所述双基因表达载体为上述的双基因组与含有双35s启动子1301质粒连接构建而成。
27.第三方面,本发明实施例还提供了上述双基因表达载体在转基因植物中的应用。
28.具体的,上述在转基因植物中的应用为将上述双基因表达载体转入转化禾本科植物,使其表达对镍的抗性和富集。
29.进一步的,所述禾本科植物为水稻。
30.下面采用具体的实施例进行详细的说明:
31.本发明所用的试验材料及其来源包括:
32.野生型水稻中花11,26℃人工气候室培养,16h光照培养。大肠杆菌(escherichia coli)dh5α由上海市农业科学院生物技术研究所植物基因工程研究室保存。克隆载体pmd-18-simple t、各类限制性内切酶、taq聚合酶、连接酶、dntp、10
×
pcr buffer和dna marker购自宝生物工程大连有限公司。所有的化学试剂都从美国西格玛化学公司和上海国药化学试剂购买。abi priam big-dye terminator dna测序试剂盒购自美国应用系统公司。
33.本发明中常规的分子生物学操作具体参见《分子克隆》【molecular cloning.2nd ed.cold spring harbor laboratory press,1989】。
34.本发明所用的试剂若未明确指明,则均购自西格玛-奥德里奇(sigma-aldrich)。
35.实施例1
36.基因合成法分别合成来源于茜草科九节属植物(psychotria gabreillae)的膜铁转运蛋白基因pgireg1s和来源于香雪球的atp磷酸核糖转移酶基因alatp-prts。
37.利用基因合成法【xiong et al.,nucl acids res,2004,32:e98】克隆来源于茜草科九节属植物(psychotria gabreillae)的膜铁转运蛋白基因pgireg1s和来源于香雪球的atp磷酸核糖转移酶基因alatp-prts,在保持pgireg1s和alatp-prts基因的氨基酸序列不变的基础上,按照水稻偏爱密码重新合成编码膜铁转运蛋白基因pgireg1s和atp磷酸核糖转移酶基因alatp-prts。
38.其中,合成后,膜铁转运蛋白基因pgireg1s,其核苷酸序列如seq id no 1所示。所述pgireg1基因编码阅读框有1518bp组成编码一个501个氨基酸的蛋白质。来源于镍超积累型香雪球的atp磷酸核糖转移酶alatp-prt基因其核苷酸序列如seq id no 2所示。所述alatp-prt基因编码阅读框有1233bp组成编码一个406个氨基酸的蛋白质。
39.基因合成设计原则包括:按水稻偏爱密码子,避免基因中出现attta等polya加尾信号,避免6个或更多的连续的a+t序列,避免5个或更多的g+c序列,g+c的比例40~60%,防止内含子切割序列,减少基因内部的两级结构hairpins,避免2、3位用cg和ta双寡核苷酸(cg在植物中易造成甲基化)。
40.利用pcr分别扩增pgireg1s和alatp-prts片段,在50μl反应体系中,内侧的引物分别为1.5ng,外侧的两个引物分别为30ng,1μl kod fx taq酶(toyobo公司,日本),5μl 10
×
pcr buffer,4μl dntp,加无菌水定容至50μl。扩增条件为:94℃预热1min;94℃,30s,50℃,30s,72℃,1min。共25个循环。
41.pcr结束后,用1%琼脂糖凝胶回收,取10μl回收产物直接与pmd-18-simple t克隆载体相连,4℃连接过夜,得到dna连接产物,高效转化到dh5α感受态细胞中。
42.从dh5α转化子细胞中抽提质粒,以抽提质粒为模板进行pcr扩增,得到了一个4875bp的片段,经测序和blast比对表明pcr法合成的基因分别与pgireg1s和alatp-prts基
因的氨基酸序列完全一致。本发明合成的pgireg1s基因,其核苷酸序列如seq id no 1所示。本发明合成的alatp-prts基因,其核苷酸序列如seq id no 2所示。
43.实施例2
44.九节属植物(psychotria gabreillae)的膜铁转运蛋白基因pgireg1s和香雪球的atp磷酸核糖转移酶基因alatp-prts双基因表达载体构建;
45.分别用bamhi和saci进行双酶切pcr产物,用1%琼脂糖凝胶回收dna片段,通过t4 dna连接酶将回收的pgireg1s和alatp-prts基因片段与含有双35s启动子1301质粒连接,酶切鉴定和序列分析测定获得了含有膜铁转运蛋白基因pgireg1s和atp磷酸核糖转移酶基因alatp-prts双基因的重组质粒eh441,如图1所示。该表达载体还包含gus报告基因和带内含子卡那霉素抗性标记基因,载体如图1所示。
46.实施例3
47.农杆菌培养
48.利用电击法将上述重组质粒eh441导入根癌农杆菌eh105s中,农杆菌蘸花法转化将构建好的含pgireg1s及alatp-prts基因的农杆菌eh105s转化到水稻愈伤中【clough et al.,the plant journal,1998,16(6)::735-743】,利用50μg/ml潮霉素进行两次筛选,后进行分化,生根,进行阳性苗鉴定后,移植入大田。待苗生长至3周左右,取叶片抽提rna,进行pcr扩增目的基因,分别获得长度为1518bp和1233bp的片段,证明目的基因已成功导入水稻中。
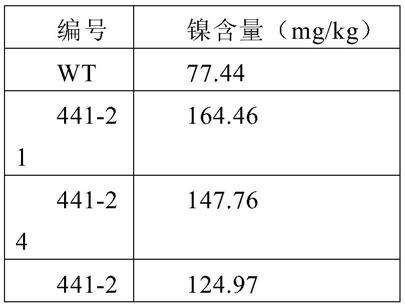
49.挑取根癌农杆菌eh105s单菌到25ml yeb培养基(50mg/l利福平)培养过夜,取5ml菌液转接到100ml yeb培养基(50mg/l利福平),培养至od600=0.7-0.8,菌液冰上放置10分钟,5000rpm离心10min,4℃,收集菌体,加入100ml无菌双蒸水清洗两次。加入4ml10%甘油悬浮菌体,转到50ml离心管。5500rpm离心10min,4℃。收集菌体,加入500μl10%甘油悬浮菌体,转到1.5ml离心管,得到农杆菌感受态细胞。取70μl上述置备好的农杆菌感受态细胞,加入1μl重组质粒eh441用去头的黄枪头混匀,转到0.1cm电击杯中。电击参数:200ω,1.7kv,2.5f,电击后立即加入800μl soc(参见《分子克隆实验指南》【sambrook and russell,2001】)培养液。培养1小时后,取100μl涂抗性板筛选转化子,28℃培养,筛选出成功导入重组质粒eh441的菌株。
50.实施例4
51.农杆菌侵染转化水稻愈伤组织
52.含目的质粒的农杆菌菌株单菌落接菌在5ml含对应抗生素的lb培养基中28℃培养2天。将5ml菌液转到500ml的液体lb培养基中28℃培养16-24小时(od=1.5-2.0),液体可以在4℃保存30天。室温下离心收集菌体,4000g离心10分钟。用等体积aam培养基悬浮。加入25mg/l的乙酰丁香酮溶液混匀后转移到三角瓶中,即得转化菌液。
53.挑选大小合适生长饱满的水稻愈伤浸入转化菌液在摇床,90转每分钟,29℃震荡培养8分钟。侵染后将愈伤用无菌滤纸吸干后,转移至含有25mg/l乙酰丁香酮的ms培养基,24℃,按暗培养2天。共培养结束后,将愈伤用无菌水洗净后吸干,转移至含有50μg/ml潮霉素、250mg/l羧苄青霉素的n6培养基进行筛选。4周后,将阳性愈伤转移至分化培养基分化成苗。
54.待苗生长至3周左右,取叶片抽提rna,反转录为cdna,利用pcr进行扩增pgireg1s
和alatp-prts片段,在50μl反应体系中,特异性引物2μg,1μl kod fx taq酶(toyobo公司,日本),5μl 10
×
pcr buffer,4μl dntp,加无菌水定容至50μl。扩增条件为:94℃预热1min;94℃,30s,50℃,30s,72℃,1min。共30个循环。
55.进行pcr扩增目的基因,结果如图2所示,图2中,m为dna mark,wt为野生型植株,1、2分别为转基因水稻。从图2中可知,野生型植株中没有检测到目的基因片段,而转入pgireg1s和alatp-prts基因植物中都有很亮的长度为280bp的条带,证明pgireg1s和alatp-prts基因已成功导入水稻并在转录水平表达。
56.实施例5
57.合成的pgireg1s和alatp-prts基因转化植物后对镍的抗性鉴定
58.将已成功转入pgireg1s及alatp-prts基因基因的转基因水稻自交纯合3代,获得纯合转化株,收取种子。播种后,25℃生长3周后,将野生型和对照分别套袋浇灌镍水溶液,使土中镍含量达500mg/kg。后期每4天浇灌80ml自来水,同时培养非转基因水稻(即野生型水稻)作为对照实验。观察转基因水稻和非转基因水稻对镍的抗性作用。浇灌一周后(野生型和转基因株系在表型上未有差异)测定野生型和转基因株系中的镍含量,结果表明,转基因植株中的镍含量明显高于野生型。浇灌3周后,野生型水稻整体生长弱于转基因株系。结果表明野生型和转基因型水稻对镍抗性差异明显,转基因水稻对镍抗性明显提高。
59.在第一次浇灌1周后进行镍含量测定,发现镍在转基因株系根与叶中的含量明显高于野生型株系,下表为浇灌镍后转pgireg1s和alatp-prts基因水稻与野生型水稻的镍含量。
[0060][0061][0062]
三周后统计野生型水稻与转基因水稻性状,野生型水稻生长株高显著低于转基因水稻,且转基因的三个株系叶大而绿,如图3所示,其中:wt为野生型水稻,441-21、441-24、441-25分别为转基因拟南芥的三个株系。实验结果表明野生型和转基因型拟南芥对镍的抗性差异明显,转基因拟南芥对镍抗性明显提高,说明本发明合成的pgireg1s和alatp-prts基因可提高植物抗和富集镍的性能。
[0063]
综上所述,本发明利用基因合成法将来源于茜草科九节属植物(psychotria gabreillae)的膜铁转运蛋白基因pgireg1s和来源于香雪球的atp磷酸核糖转移酶基因alatp-prts,进行了重新改造,得到了其核苷酸序分别如seq id no 1所示的pgireg1s基因和seq id no 2所示的alatp-prts基因。同时并将pgireg1s和alatp-prts基因成功导入水稻中,检测发现得到的转基因植株都具有很强的抗和富集镍能力,表明本发明改造合成的
pgireg1s和alatp-prts基因具有提高植物抗和富集镍的能力,可用于培育富集镍植物的技术领域。
[0064]
以上描述了本发明的基本原理、主要特征和本发明的优点。本行业的技术人员应该了解,本发明不受上述实施例的限制,上述实施例和说明书中描述的只是说明本发明的原理,在不脱离本发明精神和范围的前提下本发明还会有各种变化和改进,这些变化和改进都落入要求保护的本发明范围内。本发明要求保护范围由所附的权利要求书及其等同物界定。