arabidopsisthalianarootmorphogenesisviaauxinsignalingandethyleneinsensitive2functioning.newphytol2016,209:1496-1512.)。该物质最早从绿色木霉中分离出来,随后在多种木霉中检测到,其含量具有培养基和菌种的差异性。
4.近年来国内外文献报道表明:6pp来源于脂肪酸亚油酸的氧化,能显著抑制诸如白绢病菌(sclerotiumrolfsii)、甘薯丝核菌(rhizoctoniabataticola)、菜豆壳球孢菌(macrophominaphaseolina)、灰霉菌(botrytiscinerea)等植物病原真菌的生长。除了能抑制真菌的生长,还可以降低了串珠镰孢菌中镰孢菌酸和禾谷镰孢菌中脱氧雪腐镰孢菌烯醇的产量(wonglomp,itos,sunpapaoa:volatileorganiccompoundsemittedfromendophyticfungustrichodermaasperellumt1mediateantifungalactivity,defenseresponseandpromoteplantgrowthinlettuce(lactucasativa).fungalecology2020,43.)。此外,vinale等人研究表明6-pap对豌豆、番茄和油菜具有类似生长素的作用,即在低浓度下具有促进植物生长的作用而在较高浓度时会抑制植物生长(lees,yapm,behringerg,hungr,bennettjw:volatileorganiccompoundsemittedbytrichodermaspeciesmediateplantgrowth.fungalbiolbiotechnol2016,3:7.)。因此,6pp具有低毒性、环境相容性等特点成为开发新型生防杀菌剂优选产品。然而目前6pp主要是通过化学合成,原料价格昂贵,成本高,产率较低。如何利用微生物发酵高效合成6pp,成为开发新型生防菌剂的重要前提。
5.挥发性代谢产物(voc)作成一种理想的化学信号参与生物之间的互作,尤其在木霉菌和镰刀菌的对峙过程中起到关键的拮抗活性。目前关于6pp合成途径的遗传学研究较少,已有的研究表明:rocha-ramirez等人分析深绿木霉g蛋白亚单位tga1敲除后,影响营养菌丝生长,分生孢子形成和几丁质酶分泌,同时减少6pp和萜烯类化合物合成,但是促进代谢产物抗菌肽如trichorzianine的合成(reithnerb,brunnerk,schuhmacherr,peissli,seidlv,krskar,zeilingers:thegproteinalphasubunittga1oftrichodermaatrovirideisinvolvedinchitinaseformationanddifferentialproductionofantifungalmetabolites.fungalgenetbiol2005,42:749-760.)。无独有偶,哈茨木霉t34中转录因子thctf1缺失后,抑制了6-pentyl-2h-pyran-2-one(6-pp)及衍生物合成。reithner等人研究表明深绿木霉的促分裂原活化的蛋白激酶(mapk)δtmk1突变株合成6pp及抗菌肽类抗生素的能力增加,因此具有较高的抗真菌活性,拮抗灰霉病害(reithrierb,schuhmacherr,stoppachern,pucherm,brunnerk,zeilingers:signalingviathetrichodermaatroviridemitogen-activatedproteinkinasetmk1differentiallyaffectsmycoparasitismandplantprotection.fungalgeneticsandbiology2007,44:1123-1133.rubiomb,hermosar,reinojl,colladoig,montee:thctf1transcriptionfactoroftrichodermaharzianumisinvolvedin6-pentyl-2h-pyran-2-oneproductionandantifungalactivity.fungalgenetbiol2009,46:17-27.).6pp作为生态的抑菌剂拮抗植物病原真菌,并且在土壤和植物根际作为信号诱导植物的抗性,并促进植物生长。6-戊基-2h-吡喃-2-酮(6pp)主要由深绿木霉和哈茨木霉产生,当以1ppm低浓度剂量施用于植物表面时,它能诱导植物生长和减少疾病症状(vinalef,sivasithamparamk,ghisalbertiel,marrar,barbettimj,lih,woosl,loritom:anovelrolefortrichodermasecondarymetabolitesinthe
interactions with plants.physiological and molecular plant pathology 2008,72:80-86.)。研究表明,6-pp诱导拟南芥根生长素运输并作为乙烯依赖性反应途径的调节器 (garnica-vergara a,barrera-ortiz s,munoz-parra e,raya-gonzalez j,mendez-bravo a, macias-rodriguez l,ruiz-herrera lf,lopez-bucio j:the volatile 6-pentyl-2h-pyran-2-onefrom trichoderma atroviride regulates arabidopsis thaliana root morphogenesis via auxinsignaling and ethylene insensitive 2 functioning.new phytol 2016,209:1496-1512.)。
6.丝状真菌中velvet蛋白是调控次级代谢和性发育的重要因子。研究表明:曲霉菌(aspergillus spp.)含有vea基因编码的velvet蛋白,具有调控无性生殖和有性生殖方式转变,促进次级代谢包括聚酮类化合物和抗生素等产生(dhingra s,andes d, calvo am:vea regulates conidiation,gliotoxin production,and protease activity in theopportunistic human pathogen aspergillus fumigatus.eukaryot cell 2012,11:1531-1543.)。 bayram在自然中发表的文章表明:丝状真菌velvet蛋白是调控次级代谢和性发育的重要因子,并且受到光、温度等环境因素影响(bayram o,krappmann s,ni m,bok jw, helmstaedt k,valerius o,braus-stromeyer s,kwon nj,keller np,yu jh,braus gh: velb/vea/laea complex coordinates light signal with fungal development and secondarymetabolism.science 2008,320:1504-1506.)。2010年,mukherjee首次在绿色木霉菌中 (t.viren)克隆vea同源编码基因vel1,参与调控分生孢子和厚垣孢子形成,抑制胶霉毒素gliotoxin生物合成及其他代谢相关的基因(nrpss,pkss和甲基转移酶等)表达 (mukherjee pk,kenerley cm:regulation of morphogenesis and biocontrol properties intrichoderma virens by a velvet protein,vel1.appl environ microbiol 2010, 76:2345-2352.)。本实验室前期研究发现深绿木霉中同样有vel1基因编码velvet蛋白,具有调控菌丝形态,分生孢子和厚垣孢子形成,以及木霉菌素等次级代谢产物的功能,间接影响其生防功能。目前关于木霉菌中vel1可以负调控6pp合成尚未有报道。
技术实现要素:
7.本发明的目的在于提供一种木霉6-戊基-2h-吡喃-2-酮调控基因tavel1及其应用;具体涉及木霉6-戊基-2h-吡喃-2-酮负调控基因tavel1以及基于tavel1基因敲除株的 6pp的高产合成方法。
8.本发明的目的是通过以下技术方案来实现的:
9.第一方面,本发明涉及一种提高木霉6-戊基-2h-吡喃-2-酮产量的方法,在深绿木霉菌中敲除tavel1基因,筛选验证,获得6-戊基-2h-吡喃-2-酮高产突变株。
10.作为本发明的一个实施方案,所述敲除tavel1基因包括:构建用于敲除负调控蛋白基因tavel1的同源重组质粒;将所述重组质粒转化大肠杆菌感受态,验证、筛选阳性克隆;提取重组质粒、转化根癌农杆菌。
11.作为本发明的一个实施方案,构建同源重组质粒包括:以提取的深绿木霉基因组dna 为模板,pcr扩增,获得tavel1基因的上、下游侧翼序列;将上、下游侧翼序列分别导入敲除质粒pc1300qh的上、下游,形成重组质粒pc1300qh-δvell。
12.作为本发明的一个实施方案,扩增tavel1基因上游侧翼序列的引物为pvel1-f和
pvel1-r;pvel1-f的序列如gcgtaagcttaaatgtggctctggctcta;pvel1-r的序列如 ggtctctagagatgtcgcgtggatatgta。
13.作为本发明的一个实施方案,扩增tavel1基因下游侧翼序列的引物为dvel1-f和 dvel1-r;dvel1-f的序列如ggtaggatccctccgatgtatgagtgac;dvel1-r的序列如 cagtgagctccattatccagcagtagca。
14.作为本发明的一个实施方案,所述敲除tavel1基因包括如下步骤:
15.s1、限制性内切酶hindiii和xbaⅰ酶切目的基因上游片段pvel1和pc1300qh质粒,然后进行酶连,构建pvel1::hyg重组片段;将重组质粒pc1300qhvel1-up转化到大肠杆菌感受态中扩增并筛选阳性克隆;
16.s2、限制性内切酶bamhⅰ和sacⅰ酶切目的基因下游片段dvel1,与含有上游片段的重组质粒pc1300qhvel1-up相连,导入大肠杆菌感受态中扩增和筛选阳性克隆,提取阳性质粒即pc1300qh::pvel1::hyg::dvel1为敲除载体;
17.s3、阳性克隆菌落进行atmt转化,通过在含有潮霉素和特美汀抗性的pda平板上传代若干次后,获得稳定遗传的转化子,pcr验证结果为同源重组有条带的,即为成功敲除vel1基因的转化子t23δvell菌株。
18.作为本发明的一个实施方案,出发菌株为深绿木霉菌t23。
19.作为本发明的一个实施方案,转化根癌农杆菌是将重组质粒导入根癌农杆菌agl1 进行改良的atmt转化。
20.作为本发明的一个实施方案,所述转化采用的诱导培养基为含40umol/lmes和 200umol/las的im培养基;采用的筛选培养基为含200μg/ml潮霉素和300μg/ml 特美汀的cya培养基。
21.第二方面,本发明涉及一种深绿木霉tavel1基因在负调控木霉6-戊基-2h-吡喃-2
‑ꢀ
酮合成中的用途。
22.第三方面,本发明涉及一种深绿木霉tavel1基因敲除株在木霉6-戊基-2h-吡喃-2
‑ꢀ
酮合成中的用途。
23.第四方面,本发明涉及一种深绿木霉t23δvell菌株,所述深绿木霉t23δvell菌株为深绿木霉(trichoderma atroviride)cgmcc no.23292。
24.本发明的深绿木霉(trichoderma atroviride)t23δvell菌株,已经于2021年10月 27日递交中国微生物菌种保藏管理委员会普通微生物中心保藏,保藏地址为北京市朝阳区北辰西路1号院3号,中国科学院微生物研究所,保藏编号为cgmcc no.23292。
25.与现有技术相比,本发明具有如下有益效果:
26.1)本发明首次确定tavel1突变后,6pp合成提高的趋势,说明tavel1负调控6pp 的合成,这种转录因子负调控的情况比较少见;
27.2)本发明获得的t23δvel1菌株产生6pp的产量达200mg/l,比野生株提高了5-10 倍;
28.3)后续可通过进一步优化培养基促进6pp产量提升,为6pp的生物合成及其生防菌剂的研发奠定基础。
附图说明
29.通过阅读参照以下附图对非限制性实施例所作的详细描述,本发明的其它特征、目的和优点将会变得更明显:
30.图1为tavel1蛋白遗传背景分析示意图;
31.图2为vel1基因编码框上下游侧翼片段(a)及转化大肠杆菌转化阳性菌落克隆 (b,c),atmt遗传转化阳性克隆pcr验证(d);
32.图3为野生株t23和敲除株t23δvel1的表型比较;(a)平板表型和(b)显微孢子形态,(c)sem孢子形态观察比较,(d)光和暗条件产孢数量;
33.图4为gc-ms检测两株菌的挥发性代谢图谱,6pp为主要代谢产物;(a)gc-ms 的6pp的出峰;(b)6pp标准曲线,(c).ms的6pp离子片段及结构式;
34.图5为gc-ms检测两株菌的挥发性代谢产物含量对比;(a).发酵菌丝中6pp含量,(b).发酵液中6pp含量;
35.图6为6pp抑菌活性的体外试验结果;a.6-pp对尖孢镰孢菌抑菌率75%;b.6-pp 体外抑制番茄灰霉菌(botrytis cinerea)的试验结果。
具体实施方式
36.下面结合实施例对本发明进行详细说明。以下实施例将有助于本领域的技术人员进一步理解本发明,但不以任何形式限制本发明。应当指出的是,对本领域的普通技术人员来说,在不脱离本发明构思的前提下,还可以做出若干调整和改进。这些都属于本发明的保护范围。
37.前期实验室筛选了到一株优良6pp合成菌株深绿木霉t23,发明人通过atmt遗传转化构建并筛选获得t23菌株tavel1基因敲除株t23δvel1,该突变株的菌丝生长稀薄, 产孢和绿色色素受到抑制,同时大部分代谢产物产量下调。有意思的是,通过gc-ms 气质联用分析确定t23δvel1中在pd培养中菌丝和发酵液中6pp含量,且产量显著提高5-10倍,15d发酵液和菌丝中6pp含量达200mg/l,通过6pp抑菌试验表明100mg/l 的6pp基本完全抑制尖孢镰孢菌和灰霉菌。因此本发明的创新性在于:1.获得了深绿木霉的tavel1基因序列,并在ncbi登记,确定tavel1是负调控6pp合成的全局转录因子。2.确定了6pp合成的高产策略,即t23δvel1菌株产6pp的能力提高10倍左右,为进一步发酵产6pp提供候选优良菌株,为发展木霉菌发酵6pp为主要成分的生防菌剂提供理论依据。以下实施例提供了基于t23δvel1菌株的6pp的高产合成方法
38.实施例
39.1.材料和方法
40.1.1供试菌株及质粒和培养基
41.木霉菌菌种:深绿木霉菌(t.atroviride)t23株(accc32730),病原菌尖孢镰孢菌 (fusarium oxysporum)(saravanakumar,k.,yu,c.,dou,k.,wang,m.,li,y.,chen, j.synergistic effect of trichoderma-derived antifungal metabolites and cell wall degradingenzymes on enhanced biocontrol of fusarium oxysporum f.sp.cucumerinum.biologicalcontrol,2016,94:37-46.)和番茄灰霉菌(botrytis cinerea)(li,t.t.,zhang,j.d.,tang,j. q.,liu,z.c.,li,y.q.,chen,j.,zou,
转化技术在深绿木霉基因敲除中的应用.中国生物工程杂志,2015,35(12):58-64.),将含有目的载体的农杆菌和木霉菌孢子按照1:1进行共同培养,并在含有特美汀和潮霉素的 im平板上诱导,挑取转化子在含有特美汀和潮霉素的培养基cya上双抗筛选,挑取长出菌落的转化子于pda平板上进行传代培养和单孢分离,然后挑取性状稳定的转化子提取dna,进行pcr验证。使用δvel1-f和δvel1-r这一对引物验证敲除株,用tdna-f 和hph-r这一对引物验证过表达株,引物为dna-f和hph-r时电泳无条带,而引物为δ vel1-f和δvel1-r有条带,说明深绿木霉t23中tavel1基因敲除成功。
53.1.5木霉菌发酵液和菌丝中6pp提取
54.t23δvel1菌株和t23在pda平板活化后,取3个菌块接种到pd液体培养基,28 度180rpm摇床分别培养10、15天时,取出发酵液用4层纱布过滤分离发酵液与菌丝,发酵液用乙酸乙酯等体积萃取3次后,合并有机相,60℃下进行旋转蒸发浓缩,浓缩后加入无水硫酸钠干燥过滤,收集滤液至样品瓶待测。
55.菌丝用低温冷冻干燥,混合研磨10分钟,取0.15g冻干菌丝放置离心管,加入0.75ml 一氯甲烷与甲醇(1:1)浸泡20分钟,12000r/min低温离心10分钟,过滤取上清液,重复提取三次后混合,减压旋转蒸发,浓缩两倍,加入无水硫酸钠干燥过滤,收集滤液至样品瓶。
56.1.6 gc-ms检测6pp含量
57.待测样品通过气相色谱-质谱联用仪agilent 7890a/5975c进行检测,参数及条件设置为:气相色谱条件:色谱柱:agilent 19091s-433hp-5ms 5%phenyl methyl silox(型号:30m
×
0.25mm
×
0.25μm);进样口温:280℃;载气:高纯氦气;流速:1ml/min;升温程序:初温40℃,保持4min,以3℃/min的速率升至155℃,再以25℃/min的速率升至300℃,保持5min;进样方式:不分流进样;进样量:1μl。质谱条件:气相色谱与质谱接口温度:280℃;电离方式:电子轰击源(ei源);四级杆温度:150℃;离子源温度:230℃;电离能量:70ev;溶剂延迟:5min;质量扫描方式:全扫描(scan) 和选择离子扫描模式(sim)。
58.6pp标准液配制及标准曲线
59.取1μl 6pp纯标准品溶于10ml乙酸乙酯中,配制100ppm的6pp标准液,并以 100ppm标准液为母液,依次稀释得到50ppm、20ppm、10ppm标准液,随检测样品一同进行gc-ms分析,作为内标绘制6pp标准曲线,以进行发酵液和菌丝样品中6pp含量的测定和变化分析。
60.1.7挥发性代谢产物抑菌效果评价
61.将6pp配置成个不同浓度,分别涂布于平板上,接种待测病原菌,乙酸乙酯作为阴性对照,每个处理设3个重复。28℃下恒温培养5d,每天观察不同浓度6pp对病原菌的抑制作用。5d后用十字交叉法测量病原菌菌落直径,计算抑菌率:
[0062][0063]
其中,i为对病原菌菌落的抑菌率,c为对照组中病原菌菌落半径,t为实验组中病原菌菌落半径。
[0064]
2.结果与分析
[0065]
2.1深绿木霉tavel1基因遗传背景分析
[0066]
首先从深绿木霉克隆的tavel1基因送去测序结果为tavel1基因编码框为1693bp,
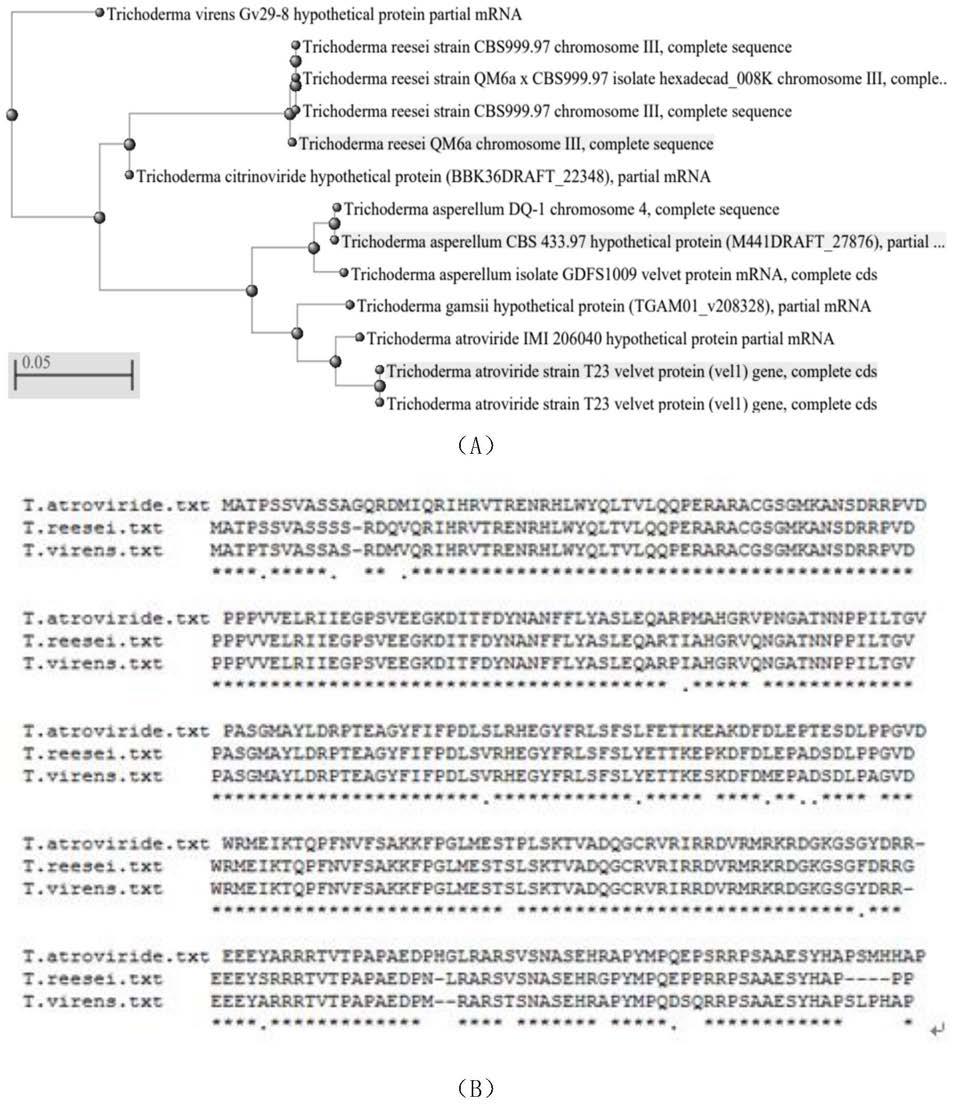
共编码545个氨基酸,ncbi的序列号kp635378,命名tavel1,genscan在线预测显示 tavel1基因共有一个内含子。根据系谱进化树和序列比对结果显示深绿木霉的tavel1蛋白与里氏木霉的同源性最高(80%的一致性),深绿木霉、里氏木霉和绿色木霉中vel1 蛋白具有高度保守性(图1)。
[0067]
2.2深绿木霉敲除载体的构建
[0068]
以深绿木霉t23野生株全基因组dna为模板,分别用pvel1-f,pvel1-r和dvel1
‑ꢀ
f,dvel1-r这两对引物扩增tavel1基因上下游侧翼序列。1%琼脂糖凝胶电泳进行分析,结果为图2所示,pcr产物长度分别是616bp和669bp,与预期一致。
[0069]
胶回收上下游侧翼片段,限制性内切酶hindiii和xbaⅰ酶切目的基因上游片段pvel1和pc1300qh质粒,然后进行酶连,酶连24h后,构建pvel1::hyg重组片段,将重组质粒pc1300qhvel1-up转化到大肠杆菌感受态e.coli5α中扩增并筛选阳性克隆。提取质粒,通过pcr和双酶切验证。限制性内切酶bamhⅰ和sacⅰ酶切下游基因片段dvel1,与含有上游片段的重组质粒pc1300qhvel1-up相连,以相同的方法导入大肠杆菌感受态中扩增和筛选阳性克隆,pcr验证后的电泳结果如所示。提取阳性克隆的质粒pc1300qh:: pvel1::hyg::dvel1即为我们所需的敲除载体。
[0070]
2.3 atmt遗传转化构建t23δvel1-8菌株
[0071]
将构建好的vel1敲除载体转化到脓杆菌agl菌株,在含有卡那霉素50mg/ml的lb 平板上培养3天后,挑取阳性克隆进行pcr验证,结果如图2所示。根据改良的atmt 转化的方法,将含有敲除载体pc1300qh::pvel1::hyg::dvel1的农杆菌(108cfu/ml)和深绿木霉t23野生型的孢子(1-8x108个孢子/ml)共培养2天后,用含有200μg/ml的潮霉素及300μg/ml的特美汀的cya作为转化子筛选培养基,筛选atmt转化子。
[0072]
将筛选培养基置于25℃培养箱培养7d左右,第一批17个转化子出现,用接种针刮取转化子边缘菌丝接种到含有200μm潮霉素和300μm特美汀的cya培养基中,28℃培养3-5天,验证其抗性。再挑取cya培养基中新生菌落边缘菌丝,置于仅含300μm特美汀的pda培养基中传代培养,一般传代3-5次,确保其抗性的遗传稳定。传代结束后,进行单孢分离,挑取由一个孢子产生的菌落边缘菌丝,再次接种到含有200μm潮霉素的抗性培养基中培养3-5天,验证其抗性已经能够稳定遗传,转化子性状稳定。
[0073]
2.4转化子的筛选和验证
[0074]
通过在含有潮霉素(200ug/ml)和特美汀(300ug/ml)抗性的pda平板上传代三次后,共获得10个稳定遗传的转化子,pcr验证结果如图3所示,t-dna插入无条带,同源重组有条带的即为成功敲除vel1基因的转化子,最终第八号转化子为所需的敲除株,标记为δvel1-8。
[0075]
2.5 vel1基因敲除株与野生株性状对比:在28℃黑暗条件下pda固体平板上培养7 天后,如图3所示,t23δvel1并未产生绿色的分生孢子,在显微镜下观察发现野生株产生大量的分生孢子,t23δvel1无任何的分生孢子,产生少量厚垣孢子,说明深绿木霉的 tavel1基因对分生孢子的产生具有促进作用,tavel1突变后,t23不产生绿色色素和分生孢子。扫描电镜结果表明突变株主要产生菌丝,且菌丝纤细,光照条件下,t23δvel1 基因敲除后在光照条件下开始产生少量孢子,说明vel1对孢子调控受光照等条件的影响。
[0076]
2.6 gc_ms检测挥发性代谢产物
[0077]
利用gc-ms检测两株菌的挥发性代谢产物,如图4,gc-ms谱图,其中36.271是目标化合物6pp,其他产物峰值都比较低对应的进一步解析度离子图;发现其中主要的挥发性代谢产物是6-吡喃酮(6pp),如图5,t23δvel1突变株总的6pp产量为在10天达到100mg/l,15天到达200mg/l,而t23产生少量6pp10-20mg/l,因此,t23δvel1中 6pp的产量是野生株的5-10倍。vel1突变影响菌丝发育和孢子形成,促进6pp合成。 6pp是一种潜在的抑制真菌病原菌的化合物,进一步检测6pp抑菌活性。
[0078]
2.7 6pp抑菌活性的体外试验
[0079]
通过6pp对镰孢菌和番茄灰霉菌的抑菌效果检测(图6),发现6pp在20ppm时就有显著抑菌能力,100ppm基本上完全抑制病原菌生长,因此构建的菌株t23δvel1具有高效合成6pp的能力,发酵10天达到100ppm。目前关于木霉菌中vel1可以负调控6pp合成尚未有报道,该菌株可以作为潜在高产6pp菌株,后期进一步通过发酵优化提高6pp产量,开发 6pp为主要成分的生物防治菌剂。
[0080]
以上对本发明的具体实施例进行了描述。需要理解的是,本发明并不局限于上述特定实施方式,本领域技术人员可以在权利要求的范围内做出各种变形或修改,这并不影响本发明的实质内容。