1.本发明涉及水果保鲜技术领域,尤其涉及一种对桑椹鲜果杀菌保鲜的方法。
背景技术:
2.桑椹味道甜美,含有丰富的营养成分,是药食两用的水果,深受广大消费者的喜爱,具有广阔的市场发展前景。然而桑椹成熟期只有一个月,处于春末夏初的5~6月份,收获温度在25~30℃,采摘时易受到机械伤害和微生物侵染。而且桑椹皮薄肉软,外表面凹凸不平,这给微生物的生长提供了良好的基地,使得桑椹非常容易腐烂变质,导致货架期极短,严重地制约了桑椹的供应和鲜果产业的发展。
3.目前,桑椹鲜果保鲜采用的方法包括低温贮藏、气调贮藏和药剂处理;其中低温贮藏和气调贮藏仅能达到抑菌效果,并不能有效杀灭致病菌;而药剂处理又不利于食品安全。因此,目前需要一种安全、有效的桑椹鲜果杀菌保鲜的方法。
技术实现要素:
4.本发明的目的在于提供一种对桑椹鲜果杀菌保鲜的方法,本发明的方法不仅安全,而且能够有效杀灭桑椹鲜果表面的致病菌。
5.为了实现上述发明目的,本发明提供以下技术方案:
6.本发明提供了一种对桑椹鲜果杀菌保鲜的方法,包括以下步骤:
7.1)将桑椹鲜果于密闭容器中单层平铺;所述密闭容器包括进气口;
8.2)通过所述进气口向密闭容器内通入大气等离子体至密闭容器内的气压为101~102kpa,进行灭菌,得到灭菌后的桑椹鲜果;所述大气等离子体的电流为2~6a,温度为9~22℃;所述灭菌的时间为30~300s。
9.优选的,所述大气等离子体的电流为2a。
10.优选的,所述大气等离子体的通入量为1~1.1m3/min。
11.优选的,在所述得到灭菌后的桑椹鲜果后,还包括对灭菌后的桑椹鲜果进行贮藏;所述贮藏的温度为1~5℃。
12.优选的,所述密闭容器的体积为8000~20000cm3。
13.优选的,所述密闭容器的规格为:长
×
宽
×
高=(40~48)cm
×
(25~33)cm
×
(8~12)cm。
14.优选的,所述密闭容器还包括出气口。
15.优选的,所述桑椹鲜果的成熟度为八成熟,果粒饱满,颜色为紫黑色。
16.本发明的有益效果:本发明提供了一种对桑椹鲜果杀菌保鲜的方法,包括以下步骤:1)将桑椹鲜果于密闭容器中单层平铺;2)向密闭容器内通入大气等离子体,进行灭菌,得到灭菌后的桑椹鲜果。本发明的方法能够显著灭活桑椹表面的细菌类、酵母和真菌,例如灰葡萄孢菌、沙门氏菌、大肠杆菌、金黄色葡萄球菌和蜡状芽孢杆菌,最高能使细菌总数减少3.65log cfu/g、酵母和霉菌总数减少1.59log cfu/g、大肠杆菌总数减少2.56log cfu/
g、金黄色葡萄球菌总数减少2.77log cfu/g、蜡状芽孢杆菌总数减少3.98log cfu/g。本发明的方法对桑椹的品质如ph、tss、硬度和色泽均没有产生显著性影响,而且能够显著地抑制桑椹的腐烂率和霉变率,最高能够降低30.00%的腐烂率和25.14%的霉变率。
附图说明
17.图1为大气低温等离子的系统搭建构架图;
18.图2为大气等离子体发生器外结构图;
19.图3为大气等离子体发生器内结构图;
20.图4为大气低温等离子体电源;
21.图5为旋涡式喷气机;
22.图6为密闭容器;
23.图7为贮藏期间桑椹细菌总数变化;
24.图8为贮藏期间桑椹酵母和霉菌总数变化;
25.图9为贮藏期间桑椹大肠杆菌的总数变化;
26.图10为贮藏期间桑椹的金黄色葡萄球菌总数变化;
27.图11为贮藏期间桑椹蜡状芽孢杆菌总数变化;
28.图12为沙门氏菌od-培养时间标准曲线;
29.图13为沙门氏菌od值-菌量标准曲线图;
30.图14-1为不同参数设置产生的大气等离子体处理后桑椹沙门氏菌的菌落数;
31.图14-2为处理当天桑椹沙门氏菌的菌落数;
32.图14-3为贮藏第2天桑椹沙门氏菌的菌落数;
33.图14-4为贮藏第4天桑椹沙门氏菌的菌落数;
34.图14-5为贮藏第8天桑椹沙门氏菌的菌落数;
35.图14-6为贮藏期间不同条件下大气等离子体处理后桑椹沙门氏菌的菌落变化;
36.图15-a为不同条件大气等离子体处理后桑椹的红绿色(a*);
37.图15-b为不同条件大气等离子体处理后桑椹的黄蓝色(b*);
38.图15-c为不同条件大气等离子体处理后桑椹的明暗度(l*);
39.图16为不同条件大气等离子体处理后桑椹的硬度;
40.图17为不同条件大气等离子体处理后桑椹的ph;
41.图18为不同条件大气等离子体处理后桑椹的可溶性固形物;
42.图19为大气等离子体处理后桑椹在20℃贮藏期间腐烂率;
43.图20为大气等离子体处理后桑椹在20℃贮藏期间霉变率。
具体实施方式
44.本发明提供了一种对桑椹鲜果杀菌保鲜的方法,包括以下步骤:
45.1)将桑椹鲜果于密闭容器中单层平铺;所述密闭容器包括进气口;
46.2)通过所述进气口向密闭容器内通入大气等离子体至密闭容器内的气压为101~102kpa,进行灭菌,得到灭菌后的桑椹鲜果;所述大气等离子体的电流为2~6a,优选为3~5a,温度为9~22℃,优选为20℃;所述灭菌的时间为30~300s,优选为50~250s,更优选为
100~150s。
47.本发明首先将桑椹鲜果于密闭容器中单层平铺;所述密闭容器包括进气口;所述进气口的规格优选为29~31mm;所述桑椹鲜果的成熟度优选为八成熟,颜色优选为紫黑色果粒饱满,采摘当天进行杀菌保鲜处理;所述桑椹鲜果的品种优选的包括大10(或称大十,无籽大十,无籽大10);所述桑椹鲜果优选为大小统一、色泽与成熟度均一致且无机械损伤的完好桑椹鲜果;所述桑椹鲜果的重量优选为5.0
±
0.50g/个;本发明所述桑椹鲜果来源于常规市售;本发明具体实施过程中,所述桑椹鲜果采摘于浙江大学紫金港校区桑树资源圃。
48.在本发明中,所述密闭容器优选的包括出气口;所述出气口的规格优选为29~31mm;所述密闭容器的体积优选为8000~20000cm3,更优选为10000~18000cm3,最优选为15000cm3;所述密闭容器的规格优选为:长
×
宽
×
高=(40~48)cm
×
(25~33)cm
×
(8~12)cm,更优选为长
×
宽
×
高=45.6cm
×
29.6cm
×
11.2cm;所述密闭容器优选的包括乐扣盒。在本发明具体实施过程中,采用乐扣盒(hpl894)作为密闭容器,尺寸为45.6cm
×
29.6cm
×
11.2cm;所述乐扣盒的两端包括两个圆孔,内径优选为30mm;所述圆孔结合垫片和螺母在圆孔处装上金属连接套管,用作进气口和出气口;所述进气口一端通过pvc管的套入与大气等离子体反应器出气腔室相连,另一端为大气等离子体出气口。
49.在本发明具体实施过程中,所述对桑椹鲜果杀菌保鲜的方法是基于大气介质阻挡放电等离子体系统(如图1所示,图1为大气低温等离子的系统搭建构架图)进行的;所述大气介质阻挡放电等离子体系统包括配电箱、大气等离子体电源、大气等离子体放电反应器、旋涡式喷气机、乐扣盒、pvc管、地线和高压线;在本发明中,所述配电箱和所述等离子体电源通过地线电连接;所述大气等离子体电源和大气等离子体放电反应器通过高压线电连接;所述配电箱和所述大气等离子体放电反应器通过地线电连接;在本发明中,所述旋涡式喷气机的喷气口和所述大气等离子体放电反应器的进气口通过pvc管连接;所述大气等离子体放电反应器的出气口和所述乐扣盒的进气口通过pvc管连接。
50.本发明中,所述大气等离子体电源、大气等离子体放电反应器、旋涡式喷气机、乐扣盒、pvc管、地线和高压线来源于常规市售。
51.本发明通过所述密闭容器的进气口向密闭容器内通入大气等离子体至密闭容器内的气压为101~102kpa,进行灭菌,得到灭菌后的桑椹鲜果;所述大气等离子体的电流为2~6a,温度为9~22℃;所述灭菌的时间为30~300s,优选为300s。本发明具体实施过程中,向密闭容器内通入大气等离子体至密闭容器内的气压为101~102kpa后,还包括采用密封膜对出气口进行密封,以防止大气等离子体漏出。在本发明中,所述大气等离子体的电流优选为2a;所述大气等离子体的通入量优选为1~1.1m3/min,更优选为1.05m3/min。
52.本发明在所述得到灭菌后的桑椹鲜果后,优选的还包括对灭菌后的桑椹鲜果进行贮藏;所述贮藏的温度优选为1~5℃,更优选为4℃。
53.下面将结合本发明中的实施例,对本发明中的技术方案进行清楚、完整地描述。显然,所描述的实施例仅仅是本发明一部分实施例,而不是全部的实施例。基于本发明中的实施例,本领域普通技术人员在没有做出创造性劳动前提下所获得的所有其他实施例,都属于本发明保护的范围。
54.本实施例所使用的装置为大气介质阻挡放电等离子体系统包括大气等离子体电源、大气等离子体放电反应器、旋涡式喷气机、乐扣盒、pvc管、高压线、地线等。大气低温等
离子的系统搭建构架图,详见图1。
55.1)反应器
56.大气等离子体反应器为单介质9管放电反应器,购自于南京苏曼等离子体科技有限公司。反应器材料均由不锈钢和聚四氟乙烯等制作。外部结构有高压接线柱、地线接线柱、观察窗和提手,高压接线柱通过高压线与电源高压接线柱相连,地线接线柱通过地线与配电箱相连。地线接线柱通过地线与配电箱相连。具体见图2,图2为大气等离子体发生器外结构图。内部结构包含:石英电极、会流条、不锈钢网、不锈钢电极、不锈钢浮动定心板、催化剂管、进气罩和出气罩,其内部构造如图3所示,图3为大气等离子体发生器内结构图。
57.等离子体产生于平行电极之间,放电宽度150mm,单边放电间隙3mm,外电极为外径内径的石英管。内电极为齿形不锈钢电极,其凹槽直径为凸台直径为腔室右侧开孔用于进气,左端开孔用于放气。放电前后两端开有玻璃窗用于观察放电及光谱诊断。本研究所使用的工作气体均为空气,通过实验室所搭旋涡式喷气机装置经过pvc管后连接放电腔室。
58.2)大气等离子体电源
59.采用数码电晕处理电源(cte-2000k),如图4所示,图4为大气低温等离子体电源。电源一侧有一个高压接线柱和一个地线接线柱,电源另一侧为操作台,可切换单介质9管反应器电源方式或液体反应器电源方式,也可控制电流,通过按键调节处理电流,调节范围为1~6a。该电源连接电压为ac 220v,功率为1500va。
60.3)旋涡式喷气机
61.采用旋涡式喷气机型号为sm-290,如图5所示,图5为旋涡式喷气机。旋涡式喷气机通过pvc管与等离子体反应器相连,将大气泵入反应器放电室。电压为220v/50hz;气压为1150mmh20;功率为290w;转速为:2900r/min;排气量为1.05m3/min。
62.4)密闭容器
63.采用乐扣盒(hpl894)作为密闭容器,尺寸为456
×
296
×
112mm,具体如图6所示,图6为密闭容器。将乐扣盒的两端钻出两个圆孔,内径为30mm,结合垫片和螺母在圆孔处装上金属连接套管,用作进气口和出气口。进气口一端通过pv管的套入与大气等离子体反应器出气腔室相连,另一端为等离子体出气口。
64.实施例1
65.1、材料
66.1)桑椹:桑椹采摘于浙江大学紫金港校区桑树资源圃,品种为大10(或称大十,无籽大十,无籽大10),成熟度为八成熟,果粒饱满,颜色为紫黑色。
67.2)主要培养基和试剂的配制:
68.平板计数琼脂培养基购自北京陆桥生物技术有限责任公司,用电子天平称取23.5g于三角瓶中,加入1000ml蒸馏水,加热搅拌溶解,于121℃高压灭菌15min,常温冷却至46℃,分装灭菌培养皿,自然凝固,4℃保存备用。
69.马铃薯葡萄糖琼脂培养基购自北京陆桥生物技术有限责任公司,用电子天平称取40.1g于三角瓶中,加入1000ml蒸馏水,加热搅拌溶解,于121℃高压灭菌15min,常温冷却至50~60℃,分装灭菌培养皿,自然凝固,4℃保存备用。
70.伊红美蓝琼脂培养基购自北京陆桥生物技术有限责任公司,用电子天平称取
37.5g于三角瓶中,加入1000ml蒸馏水,加热搅拌溶解,于121℃高压灭菌15min,常温冷却至50~60℃,分装灭菌培养皿,自然凝固,4℃保存备用。
71.甘露醇卵黄多粘菌素琼脂基础购自北京陆桥生物技术有限责任公司,用电子天平称取46.1g于三角瓶中,加入950ml蒸馏水,加热搅拌溶解,于121℃高压灭菌15min,常温冷却至55℃,每95ml加入50%卵黄液5ml及p-3e多粘菌素b溶液一支(10000iu),混匀后倾注平板,自然凝固,4℃保存备用。
72.baird-parker琼脂基础购自北京陆桥生物技术有限责任公司,用电子天平称取63g本品于三角瓶中,加入950ml蒸馏水,加热搅拌溶解,于121℃高压灭菌15min,常温冷却至55℃,每95ml加入50%亚碲酸钾卵黄增菌液5ml及baird-parker琼脂j基础95ml,混匀后倾注平板,自然凝固,4℃保存备用。
73.木糖赖氨酸脱氧胆酸钠培养基购买于北京陆桥生物技术有限责任公司,先用精准电子天平称取58.9g于三角瓶中,然后加入1000ml的蒸馏水,用玻璃棒搅拌均匀后,进行加热煮沸,常温冷却到50~60℃,分装倒入灭菌培养皿,静置片刻使其自然凝固,放入4℃冰箱保存备用。
74.氯化钠(nacl)分析纯,先用电子天平称取0.85gnacl于三角瓶中,然后加入99.15ml的蒸馏水,加热搅拌使其溶解,放入高压灭菌锅内进行121℃高压灭菌,持续15min,放入4℃低温保存备用,即为浓度为0.85%的无菌生理盐水。
75.2、方法
76.1)桑椹准备
77.挑选大小统一、色泽与成熟度均一致且无机械损伤的完好大10品种(或称大十,无籽大十,无籽大10)桑椹鲜果,数量为6,分为3组,每颗桑椹质量控制在5.0
±
0.50g,备用。
78.2)大气等离子体灭菌处理
79.4个实验组和1个ck组。不做任何处理的空白对照作为ck组,大气等离子体处理组为实验组。实验组的处理电流为2a,处理的时间是300s,对照组的处理电流为6a,处理的时间是30s。每个组设置三个样本重复,实验共重复2次。
80.3)样品原液制备
81.将灭菌处理后的桑椹放入超净工作台中,无菌取出桑椹样本(约10g),放入已灭过菌装有90ml 0.85%无菌盐水溶液的锥形瓶中,用漩涡仪振荡3min,制成1:10的样品原液。按10倍梯度逐步稀释原液,稀释到合适的3个稀释度备用(检测每种不同的菌所用的稀释度依据原始菌量选取)。
82.4)细菌总数测定
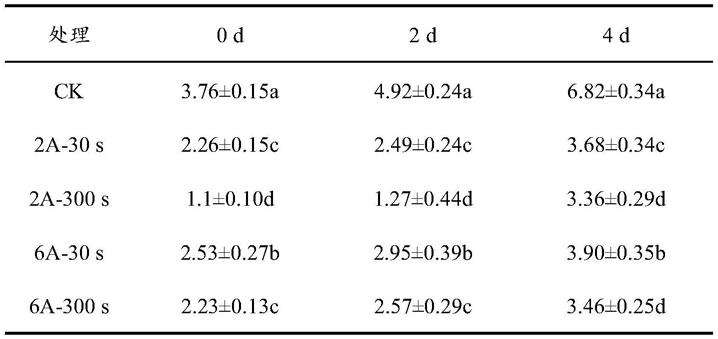
83.吸取0.1ml样品稀释液于平板计数琼脂培养基(pca)上,用一次性涂布棒将稀释液均匀涂布于pca平板,放在37℃生化培养箱中培养24~48h,以测定细菌菌落总数。等离子体杀菌处理后立即进行微生物指标检测,每48h检测1次。测定结果参见表1和图7,图7为贮藏期间桑椹细菌总数变化。
84.表1大气等离子体处理后桑椹在20℃贮藏期间的细菌总数
[0085][0086]
注:同列数值后不同字母表示具有显著性差异(p《0.05)
[0087]
由表1可知,在处理当天,大气低温等离子体处理组的细菌总数明显少于ck组的细菌总数,且均具有显著性差异。与ck组相比,2a-30s、2a-300s、6a-30s和6a-300s大气等离子体处理组能够抑制1.50log cfu/g、2.66log cfu/g、1.23log cfu/g和1.53log cfu/g的细菌。
[0088]
在处理后于20℃贮藏的第2天,大气等离子体处理组的细菌总数明显少于ck组的细菌总数,且均具有显著性差异。与对照组相比,2a-30s、2a-300s、6a-30s和6a-300s大气等离子体处理组分别能够抑制2.43logcfu/g、3.65log cfu/g、1.97log cfu/g和2.35log cfu/g的细菌。
[0089]
在20℃贮藏第4天,大气等离子体处理组的细菌总数明显少于ck组的细菌总数,且均具有显著性差异。与对照组相比,2a-30s、2a-300s、6a-30s和6a-300s大气等离子体处理组分别能够抑制3.14log cfu/g、3.46log cfu/g、2.92log cfu/g和3.36log cfu/g的细菌。
[0090]
由图7可知,贮藏0~2天期间,ck组的细菌总数上升了1.16log cfu/g,2a-30s处理组上升了0.23logcfu/g,2a-300s处理组上升了0.17log cfu/g,6a-30s处理组上升了0.42log cfu/g,6a-300s处理组上升了0.34log cfu/g。ck组细菌总数增量最大、增速最快,等离子体处理组虽然也有所上升,但上升的幅度较小,其中2a-300s处理组的细菌总数增量最少。表明大气等离子体在此贮藏期间能够发挥抑菌作用,能够抑制细菌总数的增长。
[0091]
贮藏2~4天期间,ck组的细菌总数上升了1.90log cfu/g,2a-30s处理组上升了1.19log cfu/g,2a-300s处理组上升了2.09log cfu/g,6a-30s处理组上升了0.95log cfu/g,6a-300s处理组上升了0.89log cfu/g。虽然2a-300s处理组的细菌总数相对增量最大,但其第4天的菌落数依旧是所有组中最低的,而ck组的菌落数仍然是最高的。表明在此贮藏期间,大气等离子体仍有一定抑菌作用,能够抑制细菌总数的增长。
[0092]
从处理当天到贮藏期末,ck组的细菌总数上升了3.06log cfu/g,2a-30s处理组上升了1.42log cfu/g,2a-300s处理组上升了2.26log cfu/g,6a-30s处理组上升了1.37log cfu/g,6a-300s处理组上升了1.23logcfu/g。等离子体处理组的细菌总数的上升幅度均低于ck组的细菌总数的上升幅度;不管是在第0天、第2天还是第4天,等离子体处理组的细菌总数均低于ck组的细菌总数,2a-300s处理组的细菌总数一直是处理组中最低值,说明2a-300s的等离子体处理下对细菌的抑制效果最好。
[0093]
5)酵母霉菌菌落总数测定
[0094]
吸取0.1ml样品稀释液于马铃薯葡萄糖琼脂培养基(pda)上,用一次性涂布棒将稀
释液均匀涂布于pda平板,并放在28℃生化培养箱培养5~7d,用以测定酵母霉菌菌落总数。每48h检测1次。检测结果参见表2和图8,图8为贮藏期间桑椹酵母和霉菌总数变化。
[0095]
表2大气等离子体处理后桑椹在20℃贮藏期间的酵母和霉菌总数
[0096][0097][0098]
注:表中同列数值后不同字母表示表示具有显著性差异,p《0.05
[0099]
根据表2结果可知,在处理当天,大气等离子体处理组的酵母和霉菌总数明显少于ck组的酵母和霉菌总数,且均具有显著性差异。2a-30s、2a-300s、6a-30s和6a-300s大气等离子体处理组分别能够抑制0.98logcfu/g、1.39log cfu/g、1.07log cfu/g和1.32log cfu/g的酵母和霉菌。
[0100]
在处理后于20℃贮藏的第2天,大气等离子体处理组的酵母和霉菌总数明显少于ck组的酵母和霉菌总数,且均具有显著性差异。2a-30s、2a-300s、6a-30s和6a-300s大气等离子体处理组分别能够抑制1.05logcfu/g、1.64log cfu/g、0.76log cfu/g和1.36log cfu/g的酵母和霉菌。
[0101]
在处理后于20℃贮藏的第4天,大气等离子体处理组的酵母和霉菌总数明显少于ck组的酵母和霉菌总数,且均具有显著性差异。2a-30s、2a-300s、6a-30s和6a-300s大气等离子体处理组分别能够抑制0.92log cfu/g、1.38log cfu/g、0.82log cfu/g和0.93log cfu/g的酵母和霉菌。
[0102]
由图8可知,贮藏0~2天期间,ck组、2a-30s处理组、6a-30s处理组和6a-300s处理组的酵母和霉菌总数分别上升了0.20log cfu/g、0.13logcfu/g,0.51log cfu/g和0.16log cfu/g,2a-300s处理组下降了0.05log cfu/g。6a-30s组的酵母和霉菌总数增量最大。2a-300s处理组的酵母和霉菌总数没有上升反而下降,其他组的酵母和霉菌总数均有所上升。在第2天的贮藏点,酵母和霉菌总数最高的依旧是ck组。表明大气等离子体在此贮藏期间能够发挥抑菌作用,抑制酵母和霉菌总数的增长。
[0103]
贮藏2~4天期间,所有组的酵母和霉菌总数均有所上升。ck组的酵母和霉菌总数上升了0.2log cfu/g,2a-30s、2a-300s、6a-30s和6a-300s大气等离子体处理组分别上升了0.33log cfu/g、0.46log cfu/g、0.14log cfu/g和0.63log cfu/g。等离子体增长后的酵母和霉菌总数仍然低于ck组的酵母和霉菌总数,即ck组的酵母和霉菌总数水平仍然是最高的。
[0104]
从处理当天到贮藏期末,ck组的酵母和霉菌总数上升了0.40log cfu/g,2a-30s处理组上升了0.46logcfu/g,2a-300s处理组上升了0.41log cfu/g,6a-30s处理组上升了0.65log cfu/g,6a-300s处理组上升了0.79log cfu/g。虽然等离子体处理组的酵母和霉菌
300s、6a-30s和,6a-300s的大气等离子体处理组分别上升了2.78logcfu/g、1.50log cfu/g、2.39log cfu/g和2.55log cfu/g。等离子体处理组的大肠杆菌总数上升幅度比ck组的上升幅度稍微大些,但等离子体处理组的大肠杆菌总数均低于ck组的大肠杆菌总数。不管是在第0天、第2天还是第4天,2a-300s处理组对大肠杆菌总数的抑制效果最好。
[0117]
7)金黄色葡萄球菌菌落总数测定
[0118]
吸取0.1ml样品稀释液于baird-parker琼脂培养基(bpa)上,用一次性涂布棒将稀释液均匀涂布于bpa平板,并放在生化培养箱中,于37℃培养24~48h,用以金黄色葡萄球菌菌落总数的测定。每48h检测1次。测定结果参见表4和图10,图10为贮藏期间桑椹的金黄色葡萄球菌总数变化。
[0119]
表4大气等离子体处理对桑椹在20℃贮藏期间金黄色葡萄球菌总数的影响
[0120][0121][0122]
注:表中同列数值后不同字母表示表示具有显著性差异,p《0.05
[0123]
根据表4可知,处理当天,大气等离子体处理组的金黄色葡萄球菌总数显著低于ck组的金黄色葡萄球菌总数。2a-30s、2a-300s、6a-30s和6a-300s的大气等离子体处理组分别能够抑制0.58log cfu/g、2.17logcfu/g、0.58logcfu/g和1.5log cfu/g的金黄色葡萄球菌。
[0124]
在处理后于20℃贮藏的第2天,大气等离子体处理组的金黄色葡萄球菌总数显著低于ck组的金黄色葡萄球菌总数。2a-30s、2a-300s、6a-30s和6a-300s的大气等离子体处理组分别能够抑制2.08log cfu/g、3.37logcfu/g、1.98log cfu/g和2.04log cfu/g的金黄色葡萄球菌。
[0125]
在处理后于20℃贮藏的第4天,大气等离子体处理组的金黄色葡萄球菌总数明显少于ck组的金黄色葡萄球菌总数,且均具有显著性差异。2a-30s处理组能够抑制1.70log cfu/g的金黄色葡萄球菌;2a-300s处理组能够抑制2.27log cfu/g的金黄色葡萄球菌;6a-30s处理组能够抑制1.60log cfu/g的金黄色葡萄球菌;6a-300s处理组能够抑制2.11log cfu/g的金黄色葡萄球菌。
[0126]
由图10可知,贮藏0~2天期间,ck组的金黄色葡萄球菌总数上升了1.2log cfu/g,2a-30s处理组下降了0.30log cfu/g,2a-300s处理组没有变化,均在检测水平以下,6a-30s处理组下降了0.20log cfu/g,6a-300s处理组上升了0.66log cfu/g。6a-300s组的的金黄色葡萄球菌总数增量最大,2a-30s和6a-30s处理组的金黄色葡萄球菌总数没有上升反而下降,2a-300s的在此贮藏期间均未检测到金黄色葡萄球菌。在第2天的贮藏点,金黄色葡萄球菌总数最高的依旧是ck组,且远远高于等离子体处理组。表明大气等离子体在此贮藏期间
能够发挥抑菌作用,抑制金黄色葡萄球菌总数的增长。
[0127]
贮藏2~4天期间,所有组的金黄色葡萄球菌总数均有较大幅度的增长,ck组的金黄色葡萄球菌总数上升了0.03log cfu/g,2a-30s处理组上升了0.41log cfu/g,2a-300s处理组上升了1.13log cfu/g,6a-30s处理组上升了0.41log cfu/g,6a-300s处理组下降了0.04log cfu/g。在此期间,几乎所有的处理组都呈现大致不变和上升趋势,2a-30s处理组增长值最大,但总值仍是所有组中的最低值,而ck组的金黄色葡萄球菌总数水平仍然是最高的。
[0128]
从处理当天到贮藏期末,ck组的金黄色葡萄球菌总数上升了1.23log cfu/g,2a-30s处理组上升了0.11logcfu/g,2a-300s处理组上升了1.13log cfu/g,6a-30s处理组上升了0.21log cfu/g,6a-300s处理组上升了0.62log cfu/g。等离子体处理组的金黄色葡萄球菌总数均低于ck组的的金黄色葡萄球菌总数,且具有显著性差异;不管是在第0天、第2天还是第4天,2a-300s处理组对金黄色葡萄球菌的抑制效果最好。
[0129]
8)蜡状芽孢杆菌菌落总数测定
[0130]
吸取0.1ml样品稀释液于甘露醇卵黄多粘菌素琼脂培养基(myp)上,用一次性涂布棒将稀释液均匀涂布于myp平板,并放在37℃生化培养箱中培养24~48h,测定蜡状芽孢杆菌菌落总数。每48h检测1次。结果参见表5和图11,图11为贮藏期间桑椹蜡状芽孢杆菌总数变化。
[0131]
表5大气等离子体处理对桑椹在20℃贮藏期间蜡状芽孢杆菌总数的影响
[0132][0133]
注:表中同列数值后不同字母表示表示具有显著性差异,p《0.05
[0134]
根据表5可知,在处理当天,大气等离子体处理组的蜡状芽孢杆菌总数显著低于ck组的蜡状芽孢杆菌总数。2a-30s、2a-300s、6a-30s和6a-300s的大气等离子体处理组分别能够抑制1.37log cfu/g、3.48log cfu/g、0.95logcfu/g和1.05log cfu/g的蜡状芽孢杆菌。
[0135]
在处理后于20℃贮藏的第2天,大气等离子体处理组的蜡状芽孢杆菌总数显著少于ck组的蜡状芽孢杆菌总数。2a-30s、2a-300s、6a-30s和6a-300s的大气等离子体处理组分别能够抑制2.65log cfu/g、3.98logcfu/g、0.69log cfu/g和1.63log cfu/g的蜡状芽孢杆菌。
[0136]
在处理后于20℃贮藏的第4天,大气等离子体处理组的蜡状芽孢杆菌总数明显少于ck组的蜡状芽孢杆菌总数,且均具有显著性差异。2a-30s、2a-300s、6a-30s和的6a-300s的大气等离子体分别能够抑制0.78logcfu/g、1.14log cfu/g、0.44log cfu/g和0.86log cfu/g的蜡状芽孢杆菌。
[0137]
由图11可知,贮藏0~2天期间,ck组的蜡状芽孢杆菌总数上升了0.50log cfu/g,2a-30s处理组下降了0.78log cfu/g,2a-300s处理组没有变化,未在检测水平以上,6a-30s处理组上升了0.76log cfu/g,6a-300s处理组下降了0.08log cfu/g。6a-30s组的的蜡状芽孢杆菌总数增量最大,2a-30s处理组的的蜡状芽孢杆菌总数没有上升反而下降,6a-300s和2a-300s没明显变化。在第2天的贮藏点,蜡状芽孢杆菌总数最高的依旧是ck组。表明大气等离子体在此贮藏期间能够发挥抑菌作用,抑制蜡状芽孢杆菌总数的增长。
[0138]
贮藏2~4天期间,所有组的的蜡状芽孢杆菌总数均有较大幅度的增长,ck组的蜡状芽孢杆菌总数上升了0.43log cfu/g,2a-30s处理组上升了2.30logcfu/g,2a-300s处理组上升了3.27log cfu/g,6a-30s处理组上升了0.68logcfu/g,6a-300s处理组上升了1.20log cfu/g。所有组的蜡状芽孢杆菌总数均有所增长,等离子体处理组的在此期间的增长幅度较大,而ck组的蜡状芽孢杆菌总数水平仍然是最高的。
[0139]
从处理当天到贮藏期末,ck组的蜡状芽孢杆菌总数上升了0.93log cfu/g,2a-30s处理组上升了1.52logcfu/g,2a-300s处理组上升了3.27log cfu/g,6a-30s处理组上升了1.44log cfu/g,6a-300s处理组上升了1.12log cfu/g。等离子体处理组的的蜡状芽孢杆菌总数均低于ck组的的金蜡状芽孢杆菌总数,且具有显著性差异;不管是在第0天、第2天还是第4天,2a-300s处理组对蜡状芽孢杆菌的抑制效果最好。
[0140]
实施例2大气等离子体对桑椹沙门氏菌和灰葡萄孢菌的抑菌效果
[0141]
1、材料
[0142]
1)沙门氏菌菌株选用的是常见的沙门氏菌血清型,鼠伤寒沙门氏菌(salmonellatyphimurium),由浙江大学动物科学学院预防兽医研究所实验室提供。
[0143]
2)供试冻干粉菌种灰葡萄孢atcc32762,购买于上海生物保藏中心。
[0144]
3)桑椹采摘于浙江大学紫金港校区桑树资源圃,品种为大10(或称大十,无籽大十,无籽大10)。
[0145]
4)主要培养基和试剂的配制
[0146]
脑心浸液肉汤购买于北京陆桥生物技术有限责任公司。先用精准电子天平称取52g粉末于三角瓶中,后加入1000ml的蒸馏水,加热搅拌使其溶解,并进行121℃高压灭菌,持续15min,自然冷却降温后,放入4℃保存备用。
[0147]
胰蛋白胨大豆肉汤购买于青岛海博生物技术有限公司。先用精准电子天平称取30g本品粉末于三角瓶中,加入1000ml的蒸馏水,加热搅拌使其溶解,并进行121℃高压灭菌,持续15min,自然冷却降温后,放入4℃保存备用。
[0148]
次氯酸钠(naclo)分析纯,有效氯4%,通过无菌蒸馏水的稀释,配制出浓度200ppm的次氯酸钠溶液,阴凉避光处存放备用。
[0149]
木糖赖氨酸脱氧胆酸钠培养基、马铃薯葡萄糖琼脂培养基、缓冲蛋白胨水培养基和氯化钠(nacl)分析纯的制备具体方法参考上述方案。
[0150]
2、方法
[0151]
1)沙门氏菌标准曲线的测定
[0152]
沙门氏菌的生长标准曲线图通过采用od标准曲线法进行测定。储备沙门氏菌培养物在-80℃下与50%甘油一起储存。用移液枪吸出0.2ml的沙门氏菌储备物,采用平板划线法在营养琼脂平板上进行划线,然后放在37℃生化培养箱中,培养24~48h后分离纯菌落。
挑选平板上生长良好的菌落,接种到10ml已灭菌的胰蛋白酶大豆肉汤中,进行第二次培养,放入37℃恒温摇床上进行振荡培养。在培养期间,每间隔一定的时间,取沙门氏菌培养液放入4℃冷冻高速离心机中进行离心,持续15min,过滤除去上层培养液,加入无菌生理盐水,制备为沙门氏菌悬浊液,再次进行离心,持续15min,并除去残留的培养液。利用紫外可见光分光光度计,在620nm条件下测定该时间点对应的沙门氏菌悬浊液的od值,并于同一时间将沙门氏菌悬浊液进行梯度稀释,通过平板涂布法测定具体菌落值,从而建立沙门氏菌od值-培养时间和od值-菌量的标准曲线,整个实验重复三次。结果参见图12,图12为沙门氏菌od-培养时间标准曲线。
[0153]
2)沙门氏菌的培养
[0154]
沙门氏菌培养物在-80℃下与50%甘油一起储存。用移液枪吸出0.2ml沙门氏菌的菌液,采用平板划线法在营养琼脂平板上进行划线,然后放在37℃生化培养箱中,培养24-48h后分离纯菌落。挑选生长良好的平板上的菌落,接种到10ml已灭菌的胰蛋白酶大豆肉汤中,放入37℃恒温摇床上振荡,进行第二次培养。
[0155]
根据步骤1)已测得的od-菌量标准曲线,确定需要接种量约为8log cfu/g菌悬液的od值范围和大约所需培养时间。待接近所需时间,测定菌悬液od值范围,参照标准曲线若满足log值需要,即4℃备用,实验重复三次。
[0156]
在沙门氏菌培养初期,其od值随着时间的延长而逐渐增高;培养后期,沙门氏菌od值的增速逐渐放缓。说明了沙门氏菌的生长特点,在培养初期为沙门氏菌的快速增长期,在培养后期,沙门氏菌进入了生长平台期。以沙门氏菌菌悬液的od值为自变量,以沙门氏菌计数所得的菌量对数值为因变量,绘制出沙门氏菌od值-菌量标准曲线图(图13),可知到关于沙门氏菌od值的指数方程式为y=-7.7243x2+17.324x-0.0008,其中决定系数r2=0.9984,s值为0.0900,拟合程度较好,可以用于定量计算沙门氏菌菌量。
[0157]
3)灰葡萄孢孢子悬液制备
[0158]
供试菌株灰葡萄孢冻干粉atcc32762,经过两次活化后,转接到pda培养基上进行培养,10d后,用无菌蒸馏水浸渍培养表面,获得真菌孢子。为了去除不必要的菌丝,孢子悬浮液要经过双层无菌纱布过滤。然后用血球计数板在显微镜下观察,测定悬浮液的孢子浓度。本次实验配置的孢子浓度约为106个/ml。
[0159]
4)桑椹准备
[0160]
挑选大小统一、色泽与成熟度均一致且无机械损伤的完好桑椹,每颗桑椹质量控制在5.0
±
0.50g。将挑选好的桑椹放入200ppm的次氯酸钠(naclo)中浸泡2min,以除去表面的原生自然菌群。放入生物安全柜中干燥30min,干燥15min时,对桑椹进行翻转,保证桑椹的每个面都能够被完全干燥,沙门氏菌和灰葡萄孢的桑椹准备方法一致。
[0161]
5)桑椹菌种接种
[0162]
(1)沙门氏菌的接种
[0163]
通过点接的方法,用移液枪吸出10μl培养好的沙门氏菌菌液接种到桑椹表面,然后在生物安全柜中干燥30min,等接种的沙门氏菌固定附着在桑椹表面后,进行等离子体处理。
[0164]
(2)灰葡萄孢菌的接种
[0165]
灰葡萄孢菌接种方法同沙门氏菌的一致,采用点接的方法接种灰葡萄孢孢子悬
液。
[0166]
6)大气等离子体处理
[0167]
(1)大气等离子体对桑椹沙门氏菌的处理
[0168]
沙门氏菌参数实验共设置4个实验组和1个ck组。其中接种后未处理组作为ck组,接种后大气等离子体处理组为实验组和对照组。实验组的处理电流为2a,处理的时间是300s,对照组的处理电流为6a,处理的时间是30s。每个组设置三个样本重复,实验共重复2次。
[0169]
在之后,以处理电流为2a,处理的时间是300s为基础,比较不同等离子体温度和储藏期间不同贮藏温度对桑椹表面沙门氏菌杀菌效果的影响。共设置4个实验组,2个ck组。其中不同等离子体温度分为两个水平,一个为自然产生的大气等离子体,即温度为22℃的常温大气等离子体,另一组为自然大气等离子体经过物理降温后的低温等离子体,温度为9℃。其中所采用的物理降温方法即将等离子体传送管提前在冰中预冷3h。不同贮藏温度设置了两个水平,即模拟日常的室内贮藏和低温贮藏,温度分为为20℃和4℃。
[0170]
(2)大气等离子体对桑椹灰葡萄孢菌的处理
[0171]
4个实验组和1个ck组,具体见步骤(1)。桑椹分组准备完成后,无菌转移至灭过菌的乐扣盒中,平铺一层,均匀放置。通过气管连接到大气等离子体反应器的出气口,进行大气等离子体处理。另一端为出气口。根据不同的分组,调整对应等离子体参数,进行处理。处理之后,立即用密封膜将乐扣盒的进出口层层封住。
[0172]
7)沙门氏菌计数
[0173]
将桑椹放入约30ml pw中,使用摇床以180rpm的速度洗脱桑椹果实上剩余的沙门氏菌,持续2min。将所得匀浆用0.1%pw稀释,并采用平板涂布法在xld上进行涂板,并在37℃下培养24h,进行计数,实验重复三次。测定结果参见表6、图14-1~图14-6,其中图14-1为不同参数设置产生的大气等离子体处理后桑椹沙门氏菌的菌落数;图14-2为处理当天桑椹沙门氏菌的菌落数;图14-3为贮藏第2天桑椹沙门氏菌的菌落数;图14-4为贮藏第4天桑椹沙门氏菌的菌落数;图14-5为贮藏第8天桑椹沙门氏菌的菌落数;图14-6为贮藏期间不同条件下大气等离子体处理后桑椹沙门氏菌的菌落变化。
[0174]
表6大气等离子体处理后桑椹的沙门氏菌的总数
[0175]
处理沙门氏菌总数ck5.04
±
0.06a2a-30s4.03
±
0.19b2a-300s3.42
±
0.47b6a-30s3.93
±
0.43b6a-300s3.42
±
0.40b
[0176]
图14-2~图14-6中,ck-20℃storage表示ck组常温20℃贮藏;ck4℃storage表示ck组低温4℃贮藏;roomplasma-20℃storage表示常温22℃大气等离子体处理后常温20℃贮藏;roomplasma-4℃storage表示常温22℃大气等离子体处理后低温4℃贮藏;coldplasma-20℃storage表示低温9℃大气等离子体处理后常温20℃贮藏;coldplasma-4℃storage表示低温9℃大气等离子体处理后低温4℃贮藏。
[0177]
由表6和图14-1可知,大气低温等离子体处理后,沙门氏菌的菌落数有明显的下
降,且具有显著性差异。相比未处理组,大气低温等离子体处理组均能够显著抑制沙门氏菌,其中2a-30s、2a-300s、6a-30s和6a-300s的大气等离子体处理组分别能够抑制1.01log cfu/g、1.62log cfu/g、1.11log cfu/g和1.62log cfu/g。对实验结果进行分析可知,大气等离子体处理对桑椹表面的沙门氏菌有显著的杀菌作用。随着处理时间的延长,对沙门氏菌的杀菌效果越明显,300s的等离子体处理比30s的效果好。等离子体电流大小,未对沙门氏菌的作用表现出明显趋势。2a与6a的处理没有明显的差异。效果最佳的是2a-300s处理组。
[0178]
由图14-2可知,在处理当天,相对于ck-20℃storage,roomplasma-20℃storage能够抑制1.77log cfu/g;coldplasma-20℃storage能够抑制1.44logcfu/g。相对于ck-4℃storage,roomplasma-4℃storage能够抑制1.83log cfu/g;coldplasma-4℃storage能够抑制1.37log cfu/g。在处理当天,所有等离子体处理组均能够显著降低沙门氏菌的菌落数,效果最佳的处理是roomplasma-4℃storage,能够最多减少1.83log cfu/g;效果最差的为coldplasma-20℃storage,能够最多减少1.44log cfu/g,两者抑菌效果相差0.39log cfu/g。
[0179]
由图14-3可知,在贮藏第2天,与ck-20℃storage相比,roomplasma-20℃storage能够抑制1.98log cfu/g;coldplasma-20℃storage能够抑制1.26log cfu/g。与ck-4℃storage相比,roomplasma-4℃storage能够抑制1.92log cfu/g;coldplasma-4℃storage能够抑制1.56log cfu/g。根据图3-5可知,在贮藏第2天,所有等离子体处理组均能够显著降低沙门氏菌的菌落数,效果最佳的处理是roomplasma-20℃storage,能够最多减少1.98logcfu/g;效果最差的为coldplasma-20℃storage,能够最多减少1.26log cfu/g,两者抑菌效果相差0.72log cfu/g。
[0180]
由图14-4可知,在贮藏第4天,与ck-20℃storage相比,roomplasma-20℃storage能够抑制1.83log cfu/g;coldplasma-20℃storage能够抑制1.27log cfu/gck-4℃storage相比,roomplasma-4℃storage能够抑制1.95log cfu/g;coldplasma-4℃storage能够抑制1.38log cfu/g。在贮藏第4天,所有等离子体处理组均能够显著降低沙门氏菌的菌落数,效果最佳的处理是roomplasma-4℃storage,能够最多减少1.95log cfu/g;效果最差的处理是coldplasma-20℃storage,能够最多减少1.27logcfu/g。两者抑菌效果相差0.68log cfu/g。
[0181]
由图14-5可知,在贮藏第8天,20℃贮藏的桑椹已经腐烂,因此不作检测,只测定4℃贮藏的桑椹表面的沙门氏菌。与ck-4℃storage相比,roomplasma-4℃storage能够抑制1.94log cfu/g;coldplasma-4℃storage能够抑制1.18log cfu/g。在贮藏第8天,等离子体处理组均能够显著降低沙门氏菌的菌落数,处理效果最佳的处理是roomplasma-4℃storage。处理效果最差的是coldplasma-4℃storage,两者抑菌效果相差0.76log cfu/g。
[0182]
由图14-6可知,在贮藏第0天到2天期间,ck-20℃storage的菌落数下降了0.07log cfu/g;ck-4℃storage的菌落数下降了0.28log cfu/g;room plasma-20℃storage的菌落数下降了0.28log cfu/g;roomplasma-4℃storage的菌落数下降了0.38log cfu/g;coldplasma-20℃storage的菌落数增加了0.11log cfu/g;coldplasma-4℃storage的菌落数下降了0.37log cfu/g。此阶段贮藏期,效果最佳的是roomplasma-4℃storage,能够使沙门氏菌继续下降0.38
±
0.08log cfu/g。
[0183]
在贮藏2天到4天期间,ck-20℃storage的菌落数下降了0.33log cfu/g;ck-4℃storage的菌落数下降了0.20log cfu/g;roomplasma-20℃storage的菌落数下降了0.18log cfu/g;roomplasma-4℃storage的菌落数下降了0.23logcfu/g;coldplasma-20℃storage的菌落数下降了0.34log cfu/g;coldplasma-4℃storage的菌落数下降了0.15log cfu/g。此阶段贮藏期,效果最佳的是roomplasma-4℃storage,能够使沙门氏菌继续下降0.34log cfu/g。
[0184]
在贮藏4天到8天期间,ck-4℃storage的菌落数下降了0.30log cfu/g;room plasma-4℃storage的菌落数下降了0.29log cfu/g;coldplasma-4℃storage的菌落数下降了0.08log cfu/g。此阶段贮藏期,效果最佳的是roomplasma-4℃storage。整个贮藏期间实验结果表明,相对于未处理的ck组,大气等离子体处理组均能够显著地抑制沙门氏菌的生长。在贮藏期间,大气等离子体中的长寿命粒子会继续作用,所以常温大气等离子体和低温等离子体均能够继续作用杀伤沙门氏菌,等离子体的杀菌作用仍然在持续进行。在作用效果上,常温的大气等离子体比低温的大气等离子体的杀菌效果更好。
[0185]
8)霉变率统计
[0186]
桑椹每组共计150颗用来统计霉变率,其中50颗为一个样本,共3个生物学重复。每隔2天统计一次表面霉变情况。桑椹表面出现灰霉病斑即认定为霉变。果实霉变率=霉变数/总果数。结果参见表7。
[0187]
表7大气等离子体处理后接种灰葡萄孢的桑椹的霉变率
[0188][0189][0190]
注:同列数值后不同字母表示具有显著性差异(p《0.05)
[0191]
由表7可知,在贮藏2天,2a-30s等离子体处理组能够减少54.00%的霉变;2a-300s等离子体处理组能够减少68.67%的霉变;6a-30s等离子体处理组能够减少63.34%的霉变;6a-300s等离子体处理组能够减少64.67%的霉变,处理后2天的贮藏效果最佳的是2a电流下仪器运行生成的大气低温等离子体处理桑椹300s,能够最多减少68.67%的霉变。
[0192]
处理后4天,2a-30s等离子体处理组能够减少27.33%霉变;2a-300s等离子体处理组能够减少61.33%的霉变;6a-30s等离子体处理组能够减少51.33%的霉变;6a-300s等离子体处理组能够减少58.00%的霉变,处理后4天的贮藏效果最佳的仍是2a电流下仪器运行生成的大气低温等离子体处理桑椹300s,能够最多减少61.33%的霉变率。
[0193]
在贮藏期间0天到2天的期间,所有处理组的霉变率均有增加,其中对照组的霉变率增加最多,从0增加到了78.67%,而等离子体处理组的霉变率增加范围在10.00%~
24.67%,增加最少的是2a-300s处理组。
[0194]
在贮藏期间2天到4天的期间,所有处理组的霉变率也均有增加,其中等离子体处理组的增加幅度较大,特别是2a-30s处理组增加了36.00%的霉变率,增幅最大。等离子体处理组中2a-300s的增加霉变率最少,增加了16.67%,而ck处理组的霉变率只增加了9.33%。虽然ck处理组在此阶段增加的霉变率最小,但其霉变的比例却是最高的,因为这段贮藏期的开始,ck的好果率为21.33百分制,而增加的9.33%的霉变率占所有好果率的43.74%。整个贮藏期间,大气低温等离子体能够显著抑制灰桑椹表面的葡萄孢菌的生长。
[0195]
实施例3大气等离子体对桑椹采后品质的影响
[0196]
1、材料
[0197]
桑椹于2019年5月采摘于浙江大学紫金港校区桑树资源圃,采购温度在28℃左右,品种为大10(或称大十,无籽大十,无籽大10),购买后直接运回冷库等待处理。
[0198]
2、方法
[0199]
1)桑椹准备
[0200]
挑选大小一、色泽与成熟度均一致且无机械损伤的完好桑椹,放入样品处理箱中,等待等离子体处理。
[0201]
2)大气等离子体处理
[0202]
设置1个ck组,4个等离子体处理组,其中电流两个水平分别是2a和6a,处理时间两个水平分别为30s和300s。主要包含以下分组:1)ck;2)2a-30s;3)2a-300s;4)6a-30s;5)2a-300s;每个样本需要5颗桑椹,设置三个样本重复。
[0203]
3)桑椹贮藏
[0204]
桑椹经过等离子体处理后,放入恒温智能冷库中进行贮藏,贮藏温度为20℃,湿度为90%,在处理当天,贮藏第2天和第4天进行品质测定。
[0205]
4)桑椹色泽测定
[0206]
根据cielab颜色系统对桑椹果实的色泽进行测定分析,使用前用白板进行校对。从每个处理组中随机挑选15个无腐烂、无病变且大小均匀的桑椹,使用色差仪进行色泽测定,将果实赤道处对准色差仪进行打色。记录色泽测定系统中桑椹的明暗度(l*)、红绿色(a*)和黄蓝色(b*),取平均值。测定结果参见表8、表9和表10以及图15-a、图15-b和图15-c,其中图15-a为不同条件大气等离子体处理后桑椹的红绿色(a*);图15-b为不同条件大气等离子体处理后桑椹的黄蓝色(b*);图15-c为不同条件大气等离子体处理后桑椹的明暗度(l*)。由图15-a、图15-b和图15-c的测定结果可知,不同的大气低温等离子体处理与ck组相比差异均不显著,可见大气等离子体处理对桑椹的色泽变化无显著影响(p《0.05);
[0207]
表8不同条件大气等离子体处理后桑椹的明暗度(l*)
[0208][0209]
注:表中同列数值后不同字母表示表示具有显著性差异,p《0.05
[0210]
根据表8结果可知,在第0~2天贮藏期间,所有组的l*值都无大的变化,在第2~4天贮藏期间,所有组的l*值均有所下降。贮藏第2天时,2a-30s处理组的l*值显著高于6a-300s处理组的l*值,但是所有等离子体处理组之间与ck组相比,均没有显著差异。l*值的结果表明,大气等离子体处理对桑椹的色泽没有造成显著的影响,短时间的处理更有利于桑椹保持色泽。
[0211]
表9不同条件大气等离子体处理后桑椹的红绿色(a*)
[0212][0213]
注:表中同列数值后不同字母表示表示具有显著性差异,p《0.05
[0214]
根据表9结果可知,在贮藏期间,ck和2a-30s处理组的a*值先升高后保持不变,2a-300s的处理组桑椹的a*值基本无大的变动,6a-300s和6a-300s处理桑椹的a*值逐渐缓慢下降。每一个贮藏时间点,所有组桑椹的a*值之间均没有显著性差异。
[0215]
表10不同条件大气等离子体处理后桑椹的黄蓝色(b*)
[0216]
[0217][0218]
注:表中同列数值后不同字母表示表示具有显著性差异,p《0.05
[0219]
根据表10结果可知,在贮藏期内,ck、2a-30s和6a-300s处理组的b*值先升高后保持不变,2a-300s的处理组桑椹的b*值基本无大的变动,6a-300s处理桑椹的b*值逐渐缓慢下降。每一个贮藏时间点,所有组桑椹的b*值之间均没有显著性差异。
[0220]
5)桑椹硬度测定
[0221]
使用质构仪对桑椹进行硬度测定,选用直径5.0mm的探头,探头下降深度设定为5.0mm,探头下压速率设定为1mm/s,在桑椹的赤道区域测定硬度。每个处理随机选取15个桑椹进行硬度测定,结果取平均值,单位为n.cm-2
。结果参见表11和图16,图16为不同条件大气等离子体处理后桑椹的硬度。
[0222]
硬度是用于评估水果质量最常见物理参数之一,桑椹的硬度直接反映了它的品质。根据表11可知,处理当天,桑椹的硬度范围在4.10-4.27n.cm-2
。由图15可知,ck组的硬度最低,其他等离子体处理组的硬度均比ck组的高,2a-30s和6a-30s等离子体处理组的硬度比2a-300s和6a-300s的硬度高,所有组间均无显著性差异。
[0223]
表11不同条件大气等离子体处理后桑椹的硬度
[0224][0225]
注:表中同列数值后不同字母表示表示具有显著性差异,p《0.05
[0226]
根据图16可知,在贮藏第2天,桑椹的硬度范围在3.36-3.47n.cm-2
。在第0~2天贮藏期间,所有组的硬度都有所下降,下降范围为0.72~0.81n.cm-2
。ck组的硬度仍然是最低的,2a-30s和6a-30s等离子体处理组的硬度比2a-300s和6a-300s的硬度高,所有组之间均没有显著性差异。在贮藏第4天,桑椹的硬度范围在2.65~2.90n.cm-2
。第2~4天贮藏期间,所有组的硬度都有所下降,下降范围为0.56~0.70n.cm-2
。ck组的硬度仍是最低的,所有组之间均没有显著性差异。在整个贮藏期间,随着贮藏时间的延长,桑椹硬度逐渐下降,但处理组与对照组之间均无显著性差异,即大气等离子体处理对桑椹的硬度没有影响。
[0227]
6)桑椹ph测定
[0228]
将桑椹果实放入100目的双侧滤布中进行手动挤压,通过手持式ph计对挤出来的果汁进行测定。每个处理重复3次。结果参见表12和图17,图17为不同条件大气等离子体处理后桑椹的ph。
[0229]
表12不同条件大气等离子体处理后桑椹的ph
[0230][0231]
注:表中同列数值后不同字母表示表示具有显著性差异,p《0.05
[0232]
ph值间接反映出水果的酸度,是水果口感参考的重要指标。根据表12结果可知,在等离子体处理当天,等离子体处理组的桑椹ph与ck组的ph含量相近,且所有组的ph之间均没有显著性差异。贮藏第0~2天期间,所有组的ph值均有所上升。6a-30s和6a-300s等离子体处理组的ph值上升幅度最小,ph值也最小。ck组、2a-30s和2a-300s的ph值上升幅度相近。贮藏第2天,6a-30s和和6a-300的处理组与其他组相比,能够显著地延缓桑椹ph值的升高,减缓桑椹糖分的积累。贮藏第2-4期间,ck组ph值均有所上升,其他等离子体处理组ph值均有所下降,且6a-30s和6a-300s等离子体处理组的ph值显著低于ck。而2a-30s和2a-300s等离子体处理组的ph值和ck组以及6a-30s和6a-300s处理组均没有显著性差异。
[0233]
由图17可知,整个贮藏期间2a电流条件下的大气等离子体处理组对桑椹的ph值没有影响,而6a电流条件下的大气等离子体处理组能够有效延缓桑椹的ph值的增加,减缓桑椹糖分的积累利于桑椹的保鲜。可见大气低温等离子体处理并没有对桑椹的ph产生显著性影响。
[0234]
7)桑椹可溶性固形物(tss)测定
[0235]
桑椹的可溶性固形物含量采用便携式数显糖度计进行测定,具体的操作方法与ph的测定一致。测定结果参见表13和图18,图18为不同条件大气等离子体处理后桑椹的可溶性固形物。
[0236]
表13不同条件大气等离子体处理后桑椹的可溶性固形物
[0237][0238]
注:表中同列数值后不同字母表示表示具有显著性差异,p《0.05
[0239]
可溶性固形物(tss)是水果评价的重要指标,与糖度成正比关系。由表13结果可知,在等离子体处理当天,等离子体处理组的桑椹tss与ck组的tss含量相近,且所有组的tss之间均没有显著性差异。贮藏0~2天期间,ck组的tss值有所下降,2a-30s和2a-300s的tss值几乎没有变化,而6a-30s和6a-300s的tss值则呈现上升趋势。在贮藏第2天,6a-30s的tss值是最高的,且与ck组相比,具有显著性差异。其他等离子体处理的tss值也均比ck组的高,但没有显著性差异。
[0240]
由图18可知,贮藏期间大气等离子体处理组均能够延缓桑椹的tss含量的损失,其中6a-30s处理组在贮藏第2天能够显著提高桑椹的tss含量。其他处理组的tss与ck均无显著性差异。可见大气低温等离子体处理并没有对桑椹的tss含量产生显著性影响。
[0241]
8)桑椹腐烂率测定
[0242]
桑椹每组共计150颗用来测腐烂率,其中50颗为一个样本,共3个生物学重复。桑椹处理过后,在温度为20℃、湿度为90%的紫外灭菌的恒温智能保鲜库中贮藏,每隔2天统计一次表面腐烂率情况。腐烂率分级计算,无腐烂为0级,25%以内为1级,25%~50%为2级,50%~75%为3级,75%~100%为4级。
[0243]
可根据以下公式,求出桑椹腐烂率。
[0244]
桑椹腐烂率=(1*1级对应腐烂个数)+(2*2级对应腐烂个数)+(3*3级对应腐烂个数)+(4*4级对应腐烂个数)(/4*总果数)。
[0245]
测定结果参见表14和图19。
[0246]
表14大气等离子体处理后桑椹在20℃贮藏期间腐烂率
[0247]
[0248]
注:表中同列数值后不同字母表示具有显著性差异(p《0.05)
[0249]
根据表14可知,在处理当天,所有组的腐烂率均大于30.00%,原因在于当天大气等离子体处理之后,等待了6h后才进行腐烂率的统计。相对于ck组,大气等离子体处理组均能够一定程度地降低腐烂率,但是未表现出没有显著性差异,原因是在处理当天,还未进行贮藏,腐烂率还未受到处理的影响。在贮藏期第2天,相对于ck组,2a-30s等离子体处理组能够降低18.66%的腐烂率,且具有显著性差异;2a-300s等离子体处理组能够19.55%的腐烂率,且具有显著性差异;6a-30s等离子体处理组能够15.72%的腐烂率,无显著性差异;6a-300s等离子体处理组能够18.55%的腐烂率,且具有显著性差异;在贮藏期第4天,相对于ck组,2a-30s等离子体处理组能够降低23.56%的腐烂率,且具有显著性差异;2a-300s等离子体处理组能够30.00%的腐烂率,且具有显著性差异;6a-30s等离子体处理组能够17.96%的腐烂率,且具有显著性差异;6a-300s等离子体处理组能够21.10%的腐烂率,且具有显著性差异;在贮藏期第8天,所有组桑椹的腐烂率均达到了85%以上。相对于ck组ck,2a-30s等离子体处理组能够降低1.78%的腐烂率,无显著性差异;2a-300s等离子体处理组能够6.00%的腐烂率,无显著性差异;6a-30s等离子体处理组能够3.95%的腐烂率,无显著性差异;6a-300s等离子体处理组能够11.79%的腐烂率,且具有显著性差异。
[0250]
由图19可知,整个贮藏期间,所有处理组的腐烂率均呈现上升趋势,贮藏第0~4天期间,ck组的腐烂率上升幅度最大,腐烂率最高,等离子体处理组的上升趋势相似且幅度较低。贮藏第4~8天期间,等离子体处理组的腐烂率上升幅度较大,与ck组之间的腐烂率差值比第0~4天的小。综上可知,大气低温等离子体能够显著降低桑椹在20℃贮藏4天期间的腐烂率。
[0251]
9)桑椹霉变率测定
[0252]
桑椹每组共计150颗用来测霉变率,50颗为一个样本,3个生物学重复。桑椹处理后,在温度为20℃、湿度为80℃的恒温智能保鲜库贮藏,每隔2天统计一次表面霉变率。桑椹表面出现灰霉病斑即认定为霉变,可根据以下公式求得。果实霉变率=霉变数/总果数。测定结果参见表15和图20,图20为大气等离子体处理后桑椹在20℃贮藏期间霉变率。
[0253]
表15大气等离子体处理后桑椹在20℃贮藏期间霉变率
[0254][0255]
注:同列数值后不同字母表示具有显著性差异(p《0.05)
[0256]
根据表15可知,在处理当天和贮藏期第2天,所有组均没有出现霉变情况,因致霉菌生长需要时间。贮藏4天后,与对照组相比,2a-30s、2a-300s和6a-300s大气等离子体处理组显著地降低了桑椹果实的霉变率,分别较对照组降低13.33%、13.33%和25.14%。然而,
6a-30s等离子体处理对桑椹果实的霉变率无显著性影响。
[0257]
在贮藏期第8天,所有组桑椹的霉变率均达到了35.00%以上。与对照组相比,2a-300s和6a-300s大气等离子体处理组显著地降低了桑椹果实的霉变率,分别较对照组降低26.00%和31.98%的霉变率。然而,6a-30s等离子体处理对桑椹果实的霉变率无显著性影响。根据图20可知,在贮藏第2~8天期间,所有处理组的霉变率均呈现上升趋势,其中贮藏第2~4天期间,ck组的霉变率上升幅度最大,霉变率是所有组中最高的。贮藏第4~8天期间,等离子体处理组的霉变率上升趋势与ck组相近。贮藏期末,300s处理效果比30s的效果好。综上可知,大气低温等离子体能够显著降低桑椹在20℃贮藏4天期间的腐烂率。
[0258]
以上所述仅是本发明的优选实施方式,应当指出,对于本技术领域的普通技术人员来说,在不脱离本发明原理的前提下,还可以做出若干改进和润饰,这些改进和润饰也应视为本发明的保护范围。