高等植物acs双酶活关键结构域替换制备单酶突变体及应用
技术领域
1.本发明属于基因工程技术领域,尤其涉及高等植物acs双酶活关键结构域替换制备单酶突变体及应用。
背景技术:
2.乙烯是最古老的植物激素之一,对高等植物生长发育的诸多方面,包括种子萌发、细胞伸长、根系发育、叶片伸展、开花与果实成熟、器官衰老与脱落等都起着重要的调节作用。同时,乙烯在植物对生物与非生物逆境响应与抵抗过程中的作用也不可或缺。在种子植物中,乙烯的生物合成主要经过三步重要的催化反应:第一步,甲硫氨酸在s-腺苷甲硫氨酸合酶的作用下生成s-腺苷甲硫氨酸(sam);第二步,sam在acc合酶(acc synthase,acs)的催化下发生γ位的c-s健断裂和α,γ位c-c环化生成1-氨基环丙烷-1-羧酸(acc),第三步,5
′‑
甲硫腺苷(mta)、acc在acc氧化酶(acc oxidase,aco)的作用下转化为乙烯。其中,acs催化sam形成acc是种子植物乙烯合成的限速步骤,acs也被称为乙烯合成的限速酶。
3.高等植物的acs由多基因家族编码,其成员的氨基酸序列包含7个保守域(box1-box7),属于磷酸吡哆醛(pyridoxal-5
’‑
phosphate,plp)依赖型蛋白酶的α超家族(superfamily)。除acs外,这个家族的其他亚族还包括氨基转移酶和c-s裂解酶等。模式植物拟南芥的acs家族成员共有12个,其中acs1-2,acs4-9和acs11编码乙烯合成关键酶acs,acs10和acs12编码以天冬氨酸和芳香族氨基酸为底物的氨基转移酶,而acs3是一个没有功能的假基因。长期以来,人们一直一致认为乙烯合成关键酶acs是单酶,只能催化sam形成acc,目前还没有报道对于乙烯合成关键酶acs活性的其他方面的研究。但是,申请人发现,高等植物中的乙烯合成关键酶acs,包括拟南芥atacs7,除了具有传统认知的acs活性外,还能以胱氨酸为底物催化生成铵根离子和丙酮酸(图1),表明其也具有c-s裂解酶活性。丙酮酸是糖酵解的终产物和线粒体三羧酸循环的能量底物,是维持呼吸代谢的非常重要的代谢物,在植物抵抗多种逆境胁迫中发挥重要作用。高等植物acs蛋白双酶活性的发现,使之成为促进植物发育、果实成熟以及抵抗逆境抗性两种不同而又紧密关联的生命过程的关键交叉点,对其双酶活性的平衡和调控是利用生物技术手段进行作物改造的重要靶标。
技术实现要素:
4.本发明实施例的目的在于提供高等植物acs双酶活关键结构域替换制备单酶突变体,旨在为利用生物技术手段平衡和调控高等植物acs双酶活性进而为进行作物改造提供重要靶标。
5.本发明实施例是这样实现的:
6.高等植物acs蛋白保守域box2的保守序列为f-q-d-y-h-g-[lm]-[pskl],该保守域对acs蛋白acs活性的维持至关重要。例如,如果将位于拟南芥第四号染色体13275200-13277188位置区间的acs7基因组中13275695-13275718位、或者其对应cdna序列中第396-419位编码box2结构域的24bp碱基tttcaagactaccacggtctcaaa,替换成小立碗藓三号染色
[0021]
r12-af:5
’‑
ggatccatgggtcttcctctaatgatggagagatcatcaaacaacaaca-3’[0022]
r12-ar:5
’‑
aagagtgcccaatgggttcccaggattagttatgagcactcctcggac-3’[0023]
r12-bf:5
’‑
actaatcctgggaacccattgggcactcttgtccaaaagaaggttctag-3’[0024]
r12-br:5
’‑
gcggccgctcaaaacctccttcgtcggtccatg-3’;
[0025]
步骤(2)将测序验证无误的载体,取20ng转化到bl21感受态细胞中,涂布在含有硫酸卡那霉素的lb平板上,37℃培养过夜,第二挑取单克隆;
[0026]
步骤(3)培养并收集转化后细胞,按照his-trap ff column说明书提取纯化各acs7单酶活性重组酶。
[0027]
进一步的技术方案,在所述步骤(1)中,基因合成的方法包括以下步骤:
[0028]
i.利用pcr首先对含有突变位点的上下游片段进行扩增,以r6-af与r6-ar为引物扩增出r6-a片段,以r6-br与r6-bf为引物扩增出r6-b片段;用r10-af与r10-ar扩增出r10-a片段;用r10-br与r10-bf扩增出r10-b片段;以r12-af与r12-ar扩增出r12-a片段,以r12-br与r12-bf扩增出r12-b片段;
[0029]
ii.以a、b片段互为引物通过融合pcr分别初步扩增获得r6、r10和r12目的片段,然后分别以该步骤获得的纯化产物为模板,以hifi hot start为dna聚合酶,以r6-af和r6-br、r10-af和r10-br、r12-af和r12-br为引物通过融合pcr分别获得r6、r10和r12目标产物;
[0030]
iii.将上述pcr产物纯化后,按照pmd_18-t vector cloning kit试剂盒将产物连接到pmd_18-t载体;经测序验证正确后,用bamh i和not i双酶切pmd_18-t载体和表达载体pet28a后将目标片段连接至pet28a,获得r6、r10和r12单酶活性重组酶核苷酸序列的表达载体。
[0031]
进一步的技术方案,在步骤i中,扩增的反应体系为:在50μl反应体系中,以含有野生型acs7基因的质粒为模板,10μm的引物各1μl,以hifi hot start为dna聚合酶。扩增的条件为:95℃5min;98℃20s;58℃15s;72℃2min;72℃10min;共25个循环。
[0032]
进一步的技术方案,在步骤ii中,扩增的反应体系为:在50μl反应体系中,分别根据a和b片段的浓度和长度,按照总体积15μl原则,计算a和b片段各自应加入的体积,以hifi hot start为dna聚合酶。扩增的条件为:95℃5min;98℃20s;58℃15s;72℃2min;72℃10min;共13个循环。
[0033]
进一步的技术方案,acs蛋白acs和c-s裂解酶双酶活性测定的操作步骤如下:
[0034]
a.acs活性的测定;将纯化的拟南芥acs7突变体蛋白或相应的负对照、acs反应缓冲液和s-腺苷甲硫氨酸一起孵育,用100mm hgcl2终止反应,然后加入新鲜配制的acc assay solution将生成的acc转化为乙烯,利用气象色谱仪分析计算乙烯生成量;
[0035]
b.c-s裂解酶活性的测定:将纯化的拟南芥acs7突变体蛋白或相应的负对照和底物一起孵育,反应结束后加入氯仿抽提蛋白;对于反应产物丙酮酸的测定,采用分光光度法检测丙酮酸与2,4-二硝基苯肼反应生成丙酮酸-2,4-二硝基苯腙的量;对于反应产物铵根离子的测定,则将上述抽提蛋白后的反应液抽滤,用氨基酸分析仪检测分析。
[0036]
进一步的技术方案,包括所述的重组酶r6、r10和r12序列应用于果实成熟和抗逆作物育种方面。
[0037]
本发明实施例提供的重组酶r12仅具有acs活性,而重组酶r6和r10只具有c-s裂解酶活性。本发明不仅鉴定了影响acs7高等植物acs蛋白acs和c-s裂解酶活性的重要结构域,
更为利用基因工程方法控制乙烯参与的多项生命过程比如果实成熟等提供了关键靶点。同时,acs蛋白c-s裂解酶单酶活性重组酶能够催化丙酮酸生成并参与细胞内的多种重要代谢路径,在抗逆作物分子育种中也具有广阔的应用前景和重要的经济价值。
附图说明
[0038]
图1为本发明拟南芥atacs7同时具有acs和c-s裂解酶双重活性的示意图;
[0039]
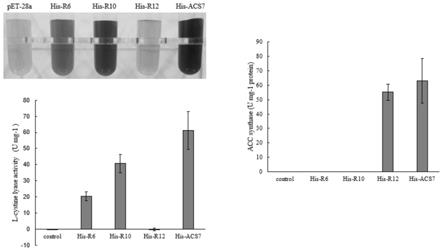
图2为本发明r6、r10重组酶只具有c-s裂解酶单酶活性,r12重组酶只有acs活性的示意图。
具体实施方式
[0040]
为了使本发明的目的、技术方案及优点更加清楚明白,以下结合附图及实施例,对本发明进行进一步详细说明。应当理解,此处所描述的具体实施例仅仅用以解释本发明,并不用于限定本发明。
[0041]
以下结合具体实施例对本发明的具体实现进行详细描述。
[0042]
首先,本发明所述三种拟南芥acs7单酶活性重组酶的制备方法,包括如下步骤:
[0043]
(1)构建含有编码所述拟南芥突变型acs7的核苷酸序列的载体
[0044]
利用基因合成的方法,合成acs7基因的上下游引物以及包含突变位点信息的引物序列如下:
[0045]
r6-af:5
’‑
ggatccatgggtcttcctctaatgatggagagatcatcaaac-3’[0046]
r6-ar:5
’‑
ctcgcctcctcggaaatttccatacaatgcgttttcac-3’[0047]
r6-bf:5
’‑
tatggaaatttccgaggaggcgagactttcagacaagcc-3’[0048]
r6-br:5
’‑
gcggccgctcaaaacctccttcgtcggt-3’[0049]
r10-af:5
’‑
ggatccatgggtcttcctctaatgatggagaga-3’[0050]
r10-ar:5
’‑
ctgagtgtcgtttgaaactgctgcaaacatgcccatccttctcgctgtccga-3’[0051]
r10-bf:5
’‑
atgggcatgtttgcagcagtttcaaacgacactcagcatatgctggcttcta-3’[0052]
r10-br:5
’‑
gcggccgctcaaaacctccttcgt-3’[0053]
r12-af:5
’‑
ggatccatgggtcttcctctaatgatggagagatcatcaaacaacaaca-3’[0054]
r12-ar:5
’‑
aagagtgcccaatgggttcccaggattagttatgagcactcctcggac-3’[0055]
r12-bf:5
’‑
actaatcctgggaacccattgggcactcttgtccaaaagaaggttctag-3’[0056]
r12-br:5
’‑
gcggccgctcaaaacctccttcgtcggtccatg-3’;
[0057]
利用pcr首先对含有突变位点的上下游片段进行扩增,以r6-af与r6-ar为引物扩增出r6-a片段,以r6-br与r6-bf为引物扩增出r6-b片段;用r10-af与r10-ar扩增出r10-a片段;用r10-br与r10-bf扩增出r10-b片段;以r12-af与r12-ar扩增出r12-a片段,以r12-br与r12-bf扩增出r12-b片段。扩增反应体系为,在50μl反应体系中,以含有野生型acs7基因的质粒为模板,10μm的引物各1μl,以hifi hot start(北京普凯瑞生物科技有限公司,货号kk2601)为dna聚合酶。扩增条件为,95℃5min;98℃20s;58℃15s;72℃2min;72℃10min;共25个循环。
[0058]
以a、b片段互为引物分别通过融合pcr分别初步扩增获得r6、r10和r12目的片段。扩增反应体系为,在50μl反应体系中,分别根据a和b片段的浓度和长度,按照总体积15μl原
则,计算a和b片段各自应加入的体积,以hifi hot start为dna聚合酶。扩增条件为,95℃5min;98℃20s;58℃15s;72℃2min;72℃10min;共13个循环。然后,分别以该步骤获得的纯化产物为模板,以hifi hot start为dna聚合酶,以r6-af和r6-br、r10-af和r10-br、r12-af和r12-br为引物通过融合pcr分别获得r6、r10和r12目标产物。
[0059]
将上述pcr产物纯化后,按照pmd_18-t vector cloning kit试剂盒将产物连接到pmd_18-t载体。经测序验证正确后,用bamh i和not i双酶切pmd_18-t载体和表达载体pet28a后将目标片段连接至pet28a,获得r6、r10和r12单酶活性重组酶核苷酸序列的表达载体。
[0060]
(2)将测序验证无误的载体,取20ng转化到bl21感受态细胞中,涂布在含有硫酸卡那霉素的lb平板上,37度培养过夜,第二挑取单克隆。
[0061]
(3)培养并收集转化后细胞,按照his-trap ff column(ge healthcare,货号17-5255-01)说明书提取纯化各acs7单酶活性重组酶。
[0062]
其次,本发明所涉及的acs蛋白acs和c-s裂解酶双酶活性测定方法如下。其中acs活性的测定方法为:将纯化的拟南芥acs7突变体蛋白或相应的负对照(转化空载体pet28a菌株的相同流程蛋白提取物)、acs反应缓冲液(50mm epps,ph8.5,10μm plp,2mm dtt)和s-腺苷甲硫氨酸(sam)一起孵育,用100mm hgcl2终止反应,然后加入新鲜配制的acc assay solution[饱和naoh:次氯酸钠=2:1(v:v)]将生成的acc转化为乙烯,利用气象色谱仪(agilent 7890a)分析计算乙烯生成量。c-s裂解酶活性测定的基本反应体系为:将纯化的拟南芥acs7突变体蛋白或相应的负对照、底物如l-胱氨酸(l-cystine)、75mm磷酸钾缓冲液ph7.6和100μm plp一起孵育,反应结束后加入氯仿抽提蛋白。对于反应产物丙酮酸的测定,采用分光光度法检测丙酮酸与2,4-二硝基苯肼反应生成丙酮酸-2,4-二硝基苯腙的量。对于反应产物铵根离子(nh
4+
)的测定,则将上述抽提蛋白后的反应液抽滤,用氨基酸分析仪(a300,membrapure gmbh)检测分析。
[0063]
如图1所示,a.氨基酸分析仪测定结果显示拟南芥atacs7可以以胱氨酸为底物催化c-s裂解反应,造成反应体系中胱氨酸含量的显著下降并生成铵根离子;b.化学显色法证明acs7可催化底物胱氨酸发生c-s键断裂生成丙酮酸。同时gc法检测证明atacs7具有正常的acs活性。his-acl1蛋白作为c-s裂解酶催化反应的正对照,以空载体pet28a为负对照(control)。
[0064]
在拟南芥中,将位于第四号染色体13275200-13277188位置区间的acs7基因组中13275695-13275718位、或其对应cdna序列中第396-419位编码box2结构域的24bp碱基tttcaagactaccacggtctcaaa,替换成小立碗藓三号染色体18432861-18432884位的24bp碱基tatggaaatttccgaggaggcgag,或将其相应mrna序列中第396-419位编码box2结构域的24bp碱基uuucaagacuaccacggucucaaa,替换成小立碗藓mrna序列相应位置的24bp碱基uauggaaauuuccgaggaggcgag,可以使acs7基因组此位置区间所编码的氨基酸由fqdyhglk变成ygnfrgge,重组后的蛋白被命名为r6。活性测定发现,与拟南芥野生型acs7蛋白相比,r6重组酶丧失acs活性,仅具有以胱氨酸为底物的c-s裂解酶活性(如图2);
[0065]
在拟南芥中,将位于第四号染色体13275200-13277188位置区间的acs7基因组中13276533-13276568位或其相应cdna序列中第1038-1073位编码box6结构域的36bp碱基atgtcgagcttcacgcttgtctcgtctcagacacaa替换成小立碗藓三号染色体18434808-18434843位
的36bp碱基atgggcatgtttgcagcagtttcaaacgacactcag,或将其相应mrna序列中第1038-1073位编码box6结构域的36bp碱基augucgagcuucacgcuugucucgucucagacacaa替换成小立碗藓mrna序列相应位置的36bp碱基augggcauguuugcagcaguuucaaacgacacucag,可以使acs7基因组此位置区间所编码的氨基酸由mssftlvssqtq变成mgmfaavsndtq,重组后的蛋白被命名为r10。活性测定发现,与拟南芥野生型acs7蛋白相比,r10重组酶丧失acs活性,仅具有以胱氨酸为底物的c-s裂解酶活性(如图2);
[0066]
在拟南芥中,将位于第四号染色体13275200-13277188位置区间的acs7基因组中13276239-13276268位或其对应cdna序列中第744-773位编码box4保守序列的30bp碱基accaacccatcgaacccattaggggcgacg替换成小立碗藓三号染色体18433956-18433985之间的30bp碱基actaatcctgggaacccattgggcactctt,或将其对应mrna序列中第744-773位编码box4保守序列的30bp碱基accaacccaucgaacccauuaggggcgacg替换成小立碗藓mrna相应序列的30bp碱基acuaauccugggaacccauugggcacucuu,可以使acs7基因组此位置区间所编码的氨基酸由tnpsnplgat变成tnpgnplgtl,重组后的蛋白被命名为r12。活性测定发现,与拟南芥野生型acs7蛋白相比,r12重组酶丧失以胱氨酸为底物的c-s裂解酶活性,仅具有acs活性(如图2)。
[0067]
本发明不仅鉴定了影响高等植物acs蛋白acs和c-s裂解酶活性的重要结构域,更为利用基因工程方法控制乙烯参与的多项生命过程比如果实成熟等提供了关键靶点,同时,重组酶r6和r10作为c-s裂解酶能够催化丙酮酸生成并参与细胞内的多种重要代谢路径,在抗逆作物分子育种中也具有广阔的应用前景和重要的经济价值。因此申请保护利用各种不同技术手段针对所有高等植物中acs同源蛋白相应于拟南芥acs7蛋白中box2、box4和box6结构域的调节和利用。
[0068]
高等植物中的乙烯合成关键酶acs,包括拟南芥atacs7,除了具有传统认知的acs活性外,还能以胱氨酸为底物催化生成铵根离子和丙酮酸,表明其也具有c-s裂解酶活性。丙酮酸是糖酵解的终产物和线粒体三羧酸循环的能量底物,是维持呼吸代谢的非常重要的代谢物,在植物抵抗多种逆境胁迫中发挥重要作用。高等植物acs蛋白双酶活性的发现,使之成为促进植物发育、果实成熟以及抵抗逆境抗性两种不同而又紧密关联的生命过程的关键交叉点,对其双酶活性的平衡和调控是利用生物技术手段进行作物改造的重要靶标。
[0069]
以上所述仅为本发明的较佳实施例而已,并不用以限制本发明,凡在本发明的精神和原则之内所作的任何修改、等同替换和改进等,均应包含在本发明的保护范围之内。
[0070]
[0071]
[0072]
[0073]
[0074]
[0075]
[0076]
[0077]