1.本发明涉及生物技术领域,尤其涉及一种分泌表达毕赤酵母内源烟酰胺核糖激酶的方法、包含毕赤酵母内源烟酰胺核糖激酶的粗酶液及其应用。
背景技术:
2.β-烟酰胺单核苷酸(β-nicotinamide mononucleotide,简称nmn)是维生素b3的吡啶-核苷形式,其中β异构体是nmn的活性形式。nmn可作为烟酰胺腺嘌呤二核苷酸(nad
+
)的直接前体物质,其生理功能是通过转化为nad+来发挥,如激活nad
+
底物依赖性酶、调节细胞存活和死亡、维持氧化还原状态等。nmn的摄入有效提高了体内nad
+
的含量,nmn可参与调节机体的内分泌并且对心脑血管疾病、神经退行性病、老化退行性疾病以及代谢性疾病等均具有较好的治疗和修复作用。
3.目前nmn主要由化学法和生物酶法合成。化学法生产nmn主要以烟酸乙酯和四乙酰核糖作为起始物质,经过缩合、脱乙酰基、磷酸化和氨解等生成nmn或者使用硅烷化试剂对烟酰胺进行硅烷化再与乙酰核糖在tmsotf的催化下进行反应生成nmn。然而化学合成反应过程冗杂且收率低,有机溶剂污染环境等问题影响了nmn的生产,使得生物法合成nmn成为主流。生物体外酶法有两条途径合成nmn,其中一条途径以烟酰胺核糖nr为底物,仅需在烟酰胺核糖激酶(nrk)和atp的作用下一步法获得nmn。另一条途径以烟酰胺和5-磷酸核糖-1-焦磷酸prpp作为底物通过多酶催化体系生成nmn。对比两条体外酶法合成途径,一步法具有转化效率高、产品纯度高等优势。
4.烟酰胺核糖激酶作为一步法转化烟酰胺核糖生成nmn的关键酶,其相关探究却非常少。现有报道的烟酰胺核糖激酶仅有酿酒酵母、人、马克斯克鲁维酵母、thermothielavioides terrestris来源,均以大肠杆菌作为表达宿主进行胞内表达,破胞处理不利于后续的纯化。2011年,k
ü
berla等人首次通过illumina测序建立的毕赤酵母cbs7435 9.35mbp高质量基因组序列,基因组序列中的注释对含烟酰胺核糖激酶在内的5007个蛋白质编码基因进行了完全分析。目前,尚无对毕赤酵母内源烟酰胺核糖激酶重组表达的相关报道。
技术实现要素:
5.针对上述问题,本发明提供了一种分泌表达毕赤酵母内源烟酰胺核糖激酶的方法、包含其的粗酶液及其应用,本发明将毕赤酵母来源的烟酰胺核酸激酶基因连接至表达载体获得重组表达载体,并导入到酵母宿主细胞,经诱导后能够高水平分泌表达烟酰胺核糖激酶。
6.本发明提供的komagataella phaffii来源烟酰胺核糖激酶基因,利用ppic9k载体分泌表达,避免了破胞处理后胞内杂蛋白对后续纯化的影响,有利于nmn的下游纯化,同时降低nmn酶法催化的生产成本,对实现nmn催化合成途径的关键酶高效表达具有较大意义。
7.本发明具体技术方案如下:
8.1.一种分泌表达毕赤酵母内源烟酰胺核糖激酶的方法,其包括:
9.将含有来源于毕赤酵母的烟酰胺核糖激酶基因的分泌型表达载体导入酵母宿主细胞中。
10.2.根据项1所述的方法,还包括:
11.培养导入了含有来源于毕赤酵母的烟酰胺核糖激酶基因的分泌型表达载体的酵母宿主细胞,进行诱导表达,
12.优选在诱导表达后直接进行离心处理以获得含有毕赤酵母内源烟酰胺核糖激酶的粗酶液。
13.3.根据项1所述的方法,其中,所述毕赤酵母内源烟酰胺核糖激酶的氨基酸序列如seq id no:2所示
14.优选所述来源于毕赤酵母的烟酰胺核糖激酶基因的核苷酸序列如seq id no:1所示。
15.4.根据项1-3中任一项所述的方法,其中,所述分泌型表达载体为ppic9、ppic9k、phil-s1、ppiczα或pyam75p,进一步优选为ppic9k。
16.5.根据项1-4中任一项所述的方法,其中,所述酵母为酿酒酵母、多形汉逊酵母、乳酸克鲁维酵母或毕赤酵母,优选为毕赤酵母,进一步优选为毕赤酵母gs115、毕赤酵母km71或毕赤酵母smd1168,进一步优选为毕赤酵母gs115。
17.6.根据项1-5中任一项所述的方法,其中,培养酵母宿主细胞的培养基为bmgy培养基,优选采用甲醇进行诱导表达。
18.7.一种包含毕赤酵母内源烟酰胺核糖激酶的粗酶液,其通过项1-6中任一项所述方法制备得到。
19.8.项7所述的包含毕赤酵母内源烟酰胺核糖激酶的粗酶液在生产烟酰胺单核苷酸中的用途。
20.9.根据项8所述的用途,其中,将包含毕赤酵母内源烟酰胺核糖激酶的粗酶液与底物接触得到烟酰胺单核苷酸,所述底物包含烟酰胺核糖和atp。
21.10.一种生产烟酰胺单核苷酸的方法,其包括:
22.将项7所述的包含毕赤酵母内源烟酰胺核糖激酶的粗酶液与底物接触得到烟酰胺单核苷酸,所述底物包含烟酰胺核糖和atp。
23.发明的效果
24.本发明将毕赤酵母来源的烟酰胺核酸激酶基因连接至表达载体,接着导入酵母宿主细胞,能够高水平表达烟酰胺核酸激酶,并且由于是分泌表达,避免了破胞处理后胞内杂蛋白对后续纯化的影响,这有利于nmn的下游纯化,同时降低nmn酶法催化的生产成本,对实现nmn催化合成途径的关键酶高效表达具有较大意义。
25.说明书附图
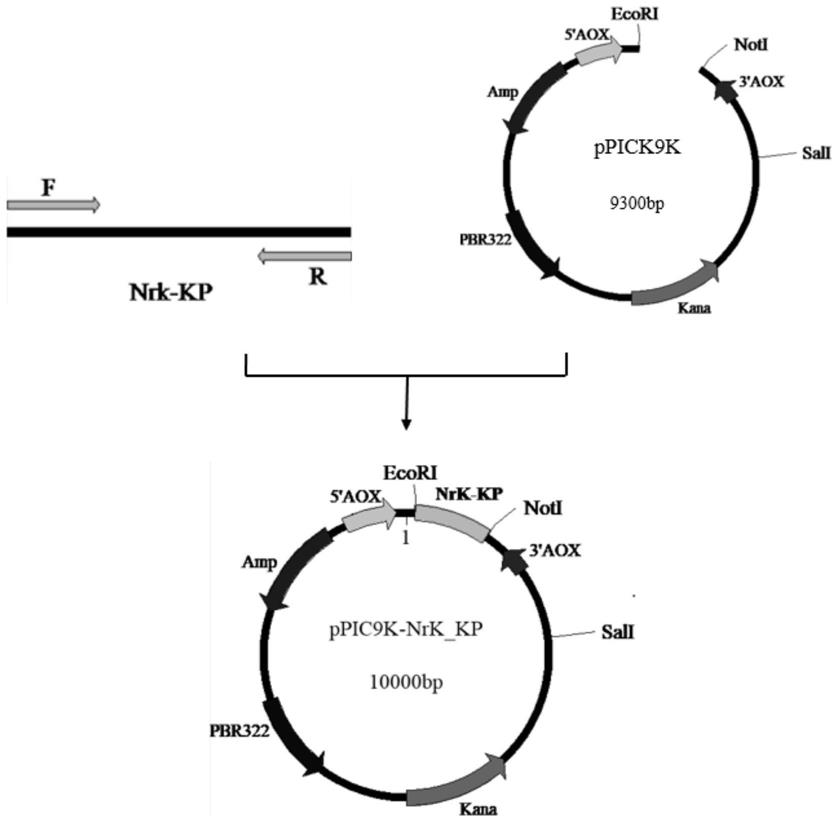
26.图1为实施例1构建重组质粒ppic9k-nrk_kp的示意图。
27.图2为重组菌烟酰胺核糖激酶发酵粗酶液sds-page蛋白电泳。其中,泳道m代表分子量为180kda的标准蛋白;泳道1代表对照菌p.pastoris gs115/ppic9k发酵粗酶液;泳道2代表komagataella phaffii来源重组菌p.pastoris gs115/ppic9k-nrk_kp发酵粗酶液;泳道3代表schizosaccharomyces pombe来源重组菌p.pastoris gs115/ppic9k-nrk_sp发酵
粗酶液;泳道4代表yarrowia lipolytica来源重组菌p.pastoris gs115/ppic9k-nrk_yl发酵粗酶液;泳道5代表wickerhamomyces ciferrii来源重组菌p.pastoris gs115/ppic9k-nrk_wc发酵粗酶液。
28.图3为实施例3中得到的烟酰胺核糖激酶产物nmn的液相图。其中,
①
为标准品的液相色谱图;
②
为对照菌p.pastoris gs115/ppic9k烟酰胺核糖激酶的产物液相色谱图;
③
为重组菌p.pastoris gs115/ppic9k-nrk_kp烟酰胺核糖激酶的产物液相色谱图。
具体实施方式
29.下面结合附图所描述的实施方式对本发明做以详细说明,其中所有附图中相同的数字表示相同的特征。虽然附图中显示了本发明的具体实施例,然而应当理解,可以以各种形式实现本发明而不应被这里阐述的实施例所限制。相反,提供这些实施例是为了能够更透彻地理解本发明,并且能够将本发明的范围完整的传达给本领域的技术人员。
30.需要说明的是,在说明书及权利要求当中使用了某些词汇来指称特定组件。本领域技术人员应可以理解,技术人员可能会用不同名词来称呼同一个组件。本说明书及权利要求并不以名词的差异作为区分组件的方式,而是以组件在功能上的差异作为区分的准则。如在通篇说明书及权利要求当中所提及的“包含”或“包括”为开放式用语,故应解释成“包含但不限定于”。说明书后续描述为实施本发明的较佳实施方式,然而所述描述乃以说明书的一般原则为目的,并非用以限定本发明的范围。本发明的保护范围当视所附权利要求所界定者为准。
31.本发明提供了一种分泌表达毕赤酵母内源烟酰胺核糖激酶nrk_kp的方法,其包括:
32.将含有来源于毕赤酵母的烟酰胺核糖激酶基因的分泌型表达载体导入酵母宿主细胞中。
33.nrk_kp指的是来源于毕赤酵母的烟酰胺核糖激酶基因所分泌表达的烟酰胺核糖激酶。
34.在一个实施方案中,所述方法还包括:
35.培养导入了含有来源于毕赤酵母的烟酰胺核糖激酶基因的分泌型表达载体的酵母宿主细胞,进行诱导表达。
36.在实验过程中,本发明的发明人意外地发现了使用毕赤酵母来源的烟酰胺核糖激酶基因连接到重组表达载体,然后导入酵母宿主细胞,能够高水平分泌表达毕赤酵母内源烟酰胺核酸激酶nrk_kp,为实现nmn的工业化生产奠定了坚实的基础。
37.在一个实施方案中,所述毕赤酵母内源烟酰胺核糖激酶nrk_kp的氨基酸序列如seq id no:2所示,优选的,所述来源于毕赤酵母的烟酰胺核糖激酶基因的核苷酸序列如seq id no:1所示,其中,seq id no:2的序列如下:
38.mtqkvviigisgssssgkttiariislvlpncllihqddfyrpneeipydpkhkaqnwdspdaidfvkfksvlhnlqndpsfvykvdslelpgddkfgadqevvrhfsklfekfsdtkfvlvdgfmmyhrgeleglfdiklmiktsystlkerrarrqgyntiggfwedppgyfdrfvwpgyynfhkdlfnepehlvkanggtlnafateqlgimafqnddggsfkqlqqdvlqslydrcitlgi
39.seq id no:1的序列如下:
40.atgacccaaa aagtggtgat aataggtatt agcggatctt ctagttccgg caaaacgacg
41.atcgcccgga taataagttt ggtcttgccc aattgcctac taattcacca agatgacttc
42.tacagaccaa acgaggaaat tccctatgat cctaaacaca aagcccaaaa ctgggactct
43.ccggatgcta tagattttgt taagtttaag tcggtactgc acaatcttca gaatgatcct
44.tcgtttgtct acaaagttga tagtttggag ttgccaggag atgataaatt tggagcagat
45.caagaggttg tgagacactt ctccaaacta ttcgaaaagt tcagtgatac aaagtttgta
46.ctagtggatg gttttatgat gtaccataga ggagagctag aaggtctctt tgacatcaaa
47.ttgatgataa agacatccta ctcaacattg aaggaaagaa gggcgagaag gcagggatat
48.aacaccattg ggggtttctg ggaagaccct ccagggtatt ttgatcgttt tgtatggcca
49.ggttactata atttccacaa agatcttttt aatgaaccag agcacttggt aaaagccaat
50.ggtggaacgc taaatgcttt tgcaacagaa caactaggaa tcatggcttt ccaaaacgat
51.gatggaggaa gtttcaaaca gctgcaacaa gacgtgcttc agtcattata tgatagatgt
52.attactttag gtatatag
53.在一个实施方案中,所述分泌型表达载体为ppic9、ppic9k、phil-s1、ppiczα或pyam75p,进一步优选为ppic9k。
54.在一个实施方案中,所述酵母为酿酒酵母、多形汉逊酵母、乳酸克鲁维酵母或毕赤酵母,优选为毕赤酵母,进一步优选为毕赤酵母gs115、毕赤酵母km71或毕赤酵母smd1168,进一步优选为毕赤酵母gs115。
55.所述多形汉逊酵母属于真菌门/子囊菌亚门/半子囊菌纲/内孢霉目/酵母科/汉逊酵母属,为单细胞低等真核生物,其为甲基营养型酵母,可在以甲醇为唯一碳源和能源的培养基中快速生长。
56.所述乳酸克鲁维酵母为克鲁维酵母属,乳酸克鲁维酵母表达外源基因既可以利用游离型载体进行表达也可以整合到基因组中进行表达,具有良好的蛋白合成和分泌能力,具备工业发酵规模的特性。
57.所述毕赤酵母是甲醇营养型酵母中的一类能够利用甲醇作为唯一碳源和能源的酵母菌,在无性生长期主要以单倍体形式存在,当环境营养限制时,常诱导2个生理类型不同的接合型单倍体细胞交配,融合成双倍体。
58.在一个实施方案中,培养酵母宿主细胞的培养基为bmgy培养基,优选的,采用甲醇进行诱导表达,优选采用含有甲醇的bmmy培养基培养以诱导表达。
59.在一个实施方案中,所述分泌表达毕赤酵母内源烟酰胺核糖激酶nrk_kp的方法包括:
60.将含有来源于毕赤酵母的烟酰胺核糖激酶基因的分泌型表达载体导入酵母宿主细胞中;
61.培养导入了含有来源于毕赤酵母的烟酰胺核糖激酶基因的分泌型表达载体的酵母宿主细胞,进行诱导表达,
62.优选在诱导表达后直接进行离心处理以获得含有毕赤酵母内源烟酰胺核糖激酶nrk_kp的粗酶液。
63.在一个实施方案中,所述分泌表达毕赤酵母内源烟酰胺核糖激酶nrk_kp的方法包括:
64.将含有来源于毕赤酵母的烟酰胺核糖激酶基因的分泌型表达载体导入酵母宿主细胞中;
65.培养酵母宿主细胞,然后进行诱导表达,
66.优选在诱导表达后直接进行离心处理以获得含有毕赤酵母内源烟酰胺核糖激酶nrk_kp的粗酶液,所述毕赤酵母内源烟酰胺核糖激酶nrk_kp的氨基酸序列如seq id no:2,
67.优选所述来源于毕赤酵母的烟酰胺核糖激酶基因的核苷酸序列如seq id no:1所示。
68.在一个实施方案中,所述分泌表达毕赤酵母内源烟酰胺核糖激酶nrk_kp的方法包括:
69.将含有来源于毕赤酵母的烟酰胺核糖激酶基因的分泌型表达载体导入酵母宿主细胞中;
70.培养酵母宿主细胞,然后进行诱导表达,
71.优选在诱导表达后直接进行离心处理以获得含有毕赤酵母内源烟酰胺核糖激酶nrk_kp的粗酶液,所述毕赤酵母内源烟酰胺核糖激酶nrk_kp的氨基酸序列如seq id no:2,
72.优选所述来源于毕赤酵母的烟酰胺核糖激酶基因的核苷酸序列如seq id no:1所示;
73.所述分泌型表达载体为ppic9、ppic9k、phil-s1、ppiczα或pyam75p,进一步优选为ppic9k。
74.在一个实施方案中,所述分泌表达毕赤酵母内源烟酰胺核糖激酶nrk_kp的方法包括:
75.将含有来源于毕赤酵母的烟酰胺核糖激酶基因的分泌型表达载体导入酵母宿主细胞中;
76.培养酵母宿主细胞,然后进行诱导表达,
77.优选在诱导表达后直接进行离心处理以获得含有毕赤酵母内源烟酰胺核糖激酶nrk_kp的粗酶液,所述毕赤酵母内源烟酰胺核糖激酶nrk_kp的氨基酸序列如seq id no:2,
78.优选所述来源于毕赤酵母的烟酰胺核糖激酶基因的核苷酸序列如seq id no:1所示;
79.所述分泌型表达载体为ppic9、ppic9k、phil-s1、ppiczα或pyam75p,进一步优选为ppic9k;
80.所述酵母为酿酒酵母、多形汉逊酵母、乳酸克鲁维酵母或毕赤酵母,优选为毕赤酵母,进一步优选为毕赤酵母gs115、毕赤酵母km71或毕赤酵母smd1168,进一步优选为毕赤酵母gs115;
81.优选的,培养酵母宿主细胞的培养基为bmgy培养基;
82.优选的,采用甲醇进行诱导表达。
83.本发明使用上述所述的方法对毕赤酵母来源的烟酰胺核糖激酶基因进行表达,其能够分泌到培养基中,这避免了破胞处理后胞内杂蛋白对后续纯化的影响,其在制备nmn时,有利于nmn的下游纯化,同时降低nmn酶法催化的生产成本。
84.本发明提供了一种包含毕赤酵母内源烟酰胺核糖激酶nrk_kp的粗酶液,其通过上述所述的方法制备得到。
85.本发明提供了上述所述的包含毕赤酵母内源烟酰胺核糖激酶nrk_kp的粗酶液在生产烟酰胺单核苷酸中的用途。
86.在一个实施方案中,将包含毕赤酵母内源烟酰胺核糖激酶nrk_kp的粗酶液与底物接触得到烟酰胺单核苷酸,所述底物包含烟酰胺核糖和atp。
87.本发明使用包含毕赤酵母内源烟酰胺核糖激酶nrk_kp的粗酶液能够高效转化烟酰胺核糖生成β-烟酰胺单核苷酸。
88.本发明提供了一种生产烟酰胺单核苷酸的方法,其包括:
89.将上述所述的包含毕赤酵母内源烟酰胺核糖激酶nrk_kp的粗酶液与底物接触得到烟酰胺单核苷酸,所述底物包含烟酰胺核糖和atp。
90.优选在37℃、ph为5.5的条件下接触。
91.实施例
92.本发明对试验中所用到的材料以及试验方法进行一般性和/或具体的描述,在下面的实施例中,如果无其他特别的说明,%表示wt%,即重量百分数。所用试剂或仪器未注明生产厂商者,均为可以通过市购获得的常规试剂产品。
93.实施例1含烟酰胺核糖激酶基因表达系统的构建
94.本发明基于ncbi数据库公开的毕赤酵母cbs 7435烟酰胺核糖激酶全长基因序列(genbank:fr839629.1),以毕赤酵母基因组为模板,pcr扩增烟酰胺核糖激酶基因核苷酸序列(其核苷酸序列如seq id no:1所示),将毕赤酵母表达载体ppic9k(购于优宝生物)用ecori和noti(购于thermo scientific)酶切、纯化后,再与烟酰胺核糖激酶基因核苷酸序列进行无缝克隆,得到重组表达载体ppic9k-nrk_kp,构建重组质粒的过程如图1所示。
95.pcr扩增体系如下,总体系50μl:100μm上游引物1.0μl,100μm下游引物1.0μl,模板1.0μl,prime star酶25μl,ddh2o 22μl。
96.pcr扩增条件如下:95℃预变性3min;95℃变性30s,55℃退火30s,72℃延伸1min,循环30次;72℃延伸5min。
97.上游引物:atgacccaaaaagtggtgataataggta(seq id no:9)
98.下游引物:ctatatacctaaagtaatacatctatcatataa(seq id no:10)
99.经dna测序比对,重组序列正确。重组表达质粒ppic9k-nrk_kp经sali快切酶线性化后电转入p.pastoris gs115表达宿主细胞中,其是在0.1cm电转杯,电容25μf,电阻200ohm,电压1kv条件下电击,重组转化子经遗传霉素g418筛选获得高拷贝重组毕赤酵母p.pastoris gs115/ppic9k-nrk_kp。
100.实施例2毕赤酵母内源的烟酰胺核糖激酶的重组表达
101.将实施例1所得到的重组工程菌p.pastoris gs115/ppic9k-nrk_kp进行摇瓶发酵培养,挑取单克隆接种于40ml的ypd培养基(酵母提取物10g/l,蛋白胨20g/l,葡萄糖20g/l),30℃、200rpm培养24h。按10%的接种量转接于40ml的初始表达培养基bmgy(酵母提取物10g/l,蛋白胨20g/l,k2hpo
4 3g/l,kh2po
4 11.8g/l,酵母基础氮源(ynb)3.4g/l,(nh4)2so
4 10g/l,生物素4
×
10-4
g/l,甘油10ml/l)中,30℃200rpm培养24h。离心收集菌体,用生理盐水洗涤菌体后更换至40ml诱导表达培养基bmmy(酵母提取物10g/l,蛋白胨20g/l,k2hpo
4 3g/l,kh2po
4 11.8g/l,ynb 3.4g/l,(nh4)2so
4 10g/l,生物素4
×
10-4
g/l,甲醇10ml/l),30℃200rpm培养,每隔24h向培养基中添加纯甲醇至终浓度为1.0%(v/v)进行诱导表达,诱导表
达96h,8000rpm 4℃下离心10min,收集发酵上清液即粗酶液,重组工程菌发酵粗酶液与对照菌(对照菌是直接将质粒电转入酵母宿主细胞)发酵粗酶液进行sds-page蛋白电泳分析,电泳分析结果如图2所示。
102.从图2可以看出,重组工程菌p.pastoris gs115/ppic9k-nrk_kp(泳道2)在烟酰胺核糖激酶理论蛋白分子量26.9kda附近有一条对应的蛋白条带。
103.对比例1其它酵母属来源的烟酰胺核糖激酶的重组表达
104.在进行上述毕赤酵母cbs7435来源的烟酰胺核糖激酶全长基因序列(genbank:fr839629.1)表达的同时,本发明的发明人还通过ncbi数据库筛选确定了其他三种不同酵母来源的烟酰胺核糖激酶基因,它们分别为schizosaccharomyces pombe来源的nrk_sp(genbank:q9c0w1.1)所述来源于毕赤酵母的烟酰胺核糖激酶基因的氨基酸序列seq id no:3所示,核苷酸序列如seq id no:4所示。
105.seq id no:3序列如下:
106.mtrktiivgvsgascsgkstlcqllhaifegsslvheddfyktdaeipvkngiadwdcqeslnldaflenlhyirdhgvlpthlrnrenknvapealieyadiikefkapaiptleqhlvfvdgfmmyvnedlinafdirlmlvtdfdtlkrrreartgyitlegfwqdpphyfenyvwpgyvhghshlfvngdvtgklldkriqlspsskmsvrdnvqwainsilnalq
107.seq id no:4序列如下:
108.atgactcgtaagacaattatcgttggcgtgtccggagcttcgtgtagtgggaagtcaacgctctgccagcttctgcacgctatttttgaaggttcctcactggttcacgaggacgatttctacaagacggatgccgagataccggtaaaaaacgggatagcggactgggattgccaagaatctctgaatttggatgcattccttgaaaacttacattatatacgagatcacggtgtcctaccgacacatctaagaaaccgggagaataagaacgtggcccccgaagctctaatcgagtacgccgatatcattaaagagtttaaagcgcctgctatccctactttggagcaacatcttgtctttgtagacgggtttatgatgtatgttaatgaagatttaataaacgcatttgacatccgccttatgttagtgaccgacttcgacaccctgaaacggagaagggaagcacgaactggctatataaccttggagggattttggcaggatccaccccattatttcgaaaattacgtctggccaggatacgtccatggtcacagccacctcttcgtgaacggcgacgtaacgggcaagctactcgacaaacgtattcaactctctccatctagtaagatgagcgtacgcgataatgttcagtgggcgattaactcgatattgaatgcgttacagtga
109.yarrowia lipolytica来源的nrk_yl(genbank:qnp96763.1)所述来源于毕赤酵母的烟酰胺核糖激酶基因的氨基酸序列seq id no:5所示,核苷酸序列如seq id no:6所示。
110.seq id no:5序列如下:
111.mirrkiaekklakeedkrevtlvalsgpsssgkstlarllrdilphviiihqddfyledsqipvidgvqdwdcpeafdfkllskvlshvkqtgelpknfkskedqnslgpaaldenavdafkrrmhpympefenklivildgimvyhdaqftelfdikilvrssyenlksrrearsgyvtlegfwkdpegyfhnivwpgylkthkqlfenenpngepskdatregirivpttdfdvaetldwvfdvildyydle
112.seq id no:6序列如下:
113.atgatccggcgcaagatagctgaaaagaagctggcaaaggaagaggataaaagagaggttacactggtagcactttctggcccttcttcctcaggtaaaagcacattggcgcgcttactgcgtgacattttgccccatgtgataatcatacaccaagacgacttctacttggaagatagccagatccctgtcatcgacggagtgcaggattgggattgcccagaagcttttgactttaaattgctatcgaaagtattaagccatgttaaacaaactggtgagcttccgaagaatttcaaa
tccaaggaggaccaaaacagtttaggccccgccgctctagatgaaaatgccgttgatgcctttaagagacggatgcacccatatatgcctgaatttgaaaacaaacttatcgtaattctcgatgggattatggtataccacgacgcgcaatttaccgaactctttgatataaaaattctcgtacgttcaagttacgaaaacttgaaatcgcggagggaggcccgatcggggtatgtcaccctggaaggcttctggaaggatccggaaggatacttccataacatagtgtggccaggttatcttaagacgcataaacagctcttcgaaaacgagaatccgaatggggagccatctaaggacgcaactcgagagggaattaggatcgtgcccacgaccgactttgatgttgcggagactctagattgggtcttcgacgtcattctagactattatgatttagagtga
114.wickerhamomyces ciferrii(genbank:xp_011277185.1)来源的nrk_wc,所述来源于毕赤酵母的烟酰胺核糖激酶基因的氨基酸序列seq id no:7所示,核苷酸序列如seq id no:8所示。
115.seq id no:7序列如下:
116.mrerkavlvaiggasssgktvsypdneipidektgeanwdcpdainfeafekeinyikqncqlspqalsyenrynsdtkdsgslsdfsddfrsqvnhitkyielencqlffidgfmlfhdpkiinlfdiklflrasyktlkerregragyqtqenfwvdppnyfdnivypayaeshkflfidqnveqdlnsrisndlgirsfannppstlssvslwaietlnlglqhlkynll
117.seq id no:8序列如下:
118.atgcgagagcgtaaggcggttctggtagcaatcggcggagctagtagtagcgggaagacagtttcctatcctgataacgaaatcccgattgatgaaaaaaccggtgaagctaactgggactgccccgacgcaattaactttgaggcgttcgagaaagaaataaactatataaaacagaattgtcagctgtcgccccaagcactttcctacgagaatagatataattctgataccaaggatagtggaagcttgtctgatttctctgacgattttagatcccaggtgaaccatattacaaaatatatagaactagaaaactgccaactgttctttattgacggctttatgctctttcatgatccaaagatcataaacctttttgatatcaagctcttcttacgtgccagctacaaaactttaaaggaacggcgggaggggagggctggttaccaaacgcaagagaatttctgggtcgaccctccaaactactttgacaatatagtataccccgcgtatgccgagtcacacaagttcttgttcatcgaccagaatgtggaacaagacctaaattccaggatttcaaatgacctaggcatccgctcatttgccaataacccgccgtctacgctctcgtcggtcagcttatgggcgattgagactcttaaccttggactccagcacttgaaatataatctactgtga
119.上述基因均根据毕赤酵母密码子偏好性对其进行了密码子优化,密码子优化的烟酰胺核糖激酶序列委托南京金斯瑞生物科技有限公司全基因合成,并克隆到毕赤酵母表达载体ppic9k的ecori和noti酶切位点之间,得到重组表达载体ppic9k-nrk_sp、ppic9k-nrk_yl和ppic9k-nrk_wc。经dna测序比对,重组序列正确。上述重组表达质粒经sali快切酶线性化后电转入p.pastoris gs115表达宿主细胞中,重组转化子经遗传霉素g418筛选获得高拷贝重组毕赤酵母,获得重组菌p.pastoris gs115/ppic9k-nrk_sp、重组菌p.pastoris gs115/ppic9k-nrk_yl、重组菌p.pastoris gs115/ppic9k-nrk_wc。三株重组菌按照实施例2的方法进行摇瓶发酵,发酵粗酶清液与对照菌发酵粗酶液进行sds-page蛋白电泳分析,电泳分析结果如图2所示。
120.从图2可以看出,仅有泳道2(实施例2)毕赤酵母内源重组菌p.pastoris gs115/ppic9k-nrk_kp在对应理论蛋白分子量26.9kda附近有一条蛋白条带,说明只有毕赤酵母来源的烟酰胺核糖激酶基因能够分泌表达,而其他来源的烟酰胺核糖激酶基因不能分泌表达。
121.实施例3:生物催化制备nmn
122.反应体系底物液中依次加入7.5mm atp,5mm mgcl2·
6h2o,3mm nr,20mm tris-hcl,ph至5.5定容至50ml。反应体系为1ml:500μl重组菌发酵粗酶液(实施例2所得到的粗酶液)或者对照发酵粗酶液分别加入到500μl上述底物液中,37℃反应30min,反应结束后立即置于沸水浴中2min,使酶失活终止反应,稀释10倍后液相检测。
123.液相检测条件如下:
124.流动相a:nah2po4;流动相b:甲醇
125.液相检测条件:波长260nm,柱温25℃,进样量20ul,流速1ml/min
126.色谱柱及仪器:使用capcell pak c18 mgⅱ柱配置安捷伦1260液相色谱仪紫外检测器梯度进样。
127.检测梯度方法:
128.时间(min)流动相a(%)流动相b(%)09914991595510851512802012.199116991
129.标准品的制备如下:
130.分别称取nmn、nr、atp和adp,用水溶解后分别加水定容于50ml容量瓶,配置成母液(nmn 0.3561g/l,atp 0.5382g/l,nr 0.5523g/l,adp 0.4764g/l),分别吸取各自母液10ml再加水定容至100ml容量瓶配置成混合标样(nmn 35.61mg/l,atp 53.82mg/l,nr 55.23mg/l,adp 47.64mg/l)
131.最终底物nr摩尔转化率可达到92.6%。体外酶法制备nmn结果的液相色谱图如图3所示,其中,
③
为重组菌p.pastoris gs115/ppic9k-nrk_kp烟酰胺核糖激酶的产物液相图,
②
为对照菌,
①
是标准品。
132.从图3可以看出,使用重组菌p.pastoris gs115/ppic9k-nrk_kp烟酰胺核糖激酶的粗酶液进行酶促反应后,可以明显看到nmn的液相峰,而对照菌
②
在对应位置无峰出现,说明所得到的粗酶液中含有烟酰胺核糖激酶,其可以催化底物反应生成nmn。
133.综上所述,本发明所述的毕赤酵母来源的烟酰胺核糖激酶基因连接到分泌型表达载体,能够高水平表达烟酰胺核酸激酶,并且表达是分泌表达,这避免了破胞处理后胞内杂蛋白对后续纯化的影响,有利于nmn的下游纯化,同时降低nmn酶法催化的生产成本,对实现nmn催化合成途径的关键酶高效表达具有较大意义。
134.以上所述,仅是本发明的较佳实施例而已,并非是对本发明作其它形式的限制,任何熟悉本专业的技术人员可能利用上述揭示的技术内容加以变更或改型为等同变化的等效实施例。但是凡是未脱离本发明技术方案内容,依据本发明的技术实质对以上实施例所作的任何简单修改、等同变化与改型,仍属于本发明技术方案的保护范围。